

Haplotypes in the dystrophin DNA segment point to a mosaic origin of modern human diversity
- PMID: 14513410
- PMCID: PMC1180505
- DOI: 10.1086/378777
Haplotypes in the dystrophin DNA segment point to a mosaic origin of modern human diversity
Abstract
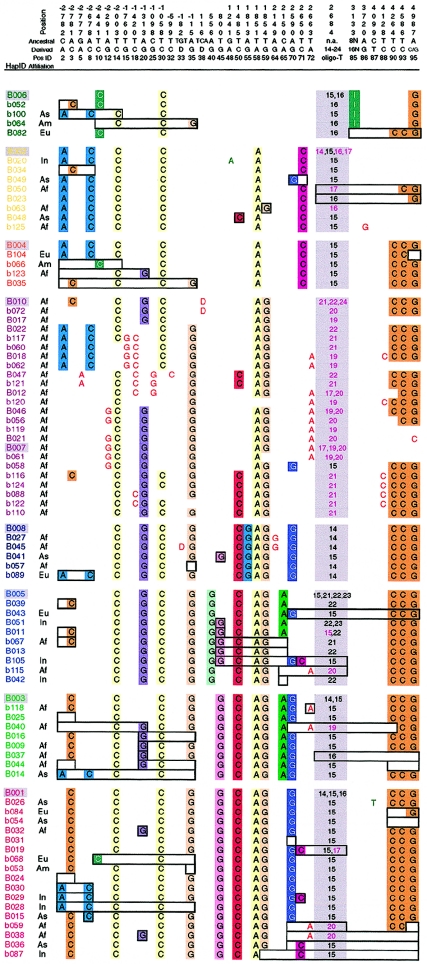
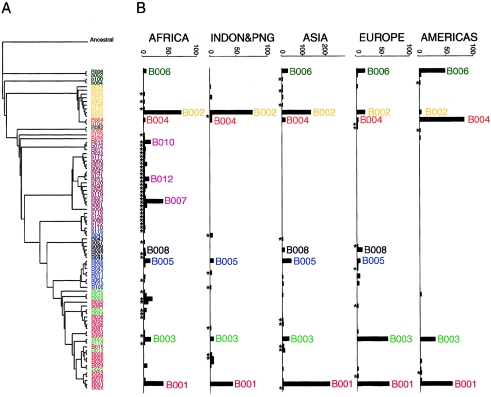
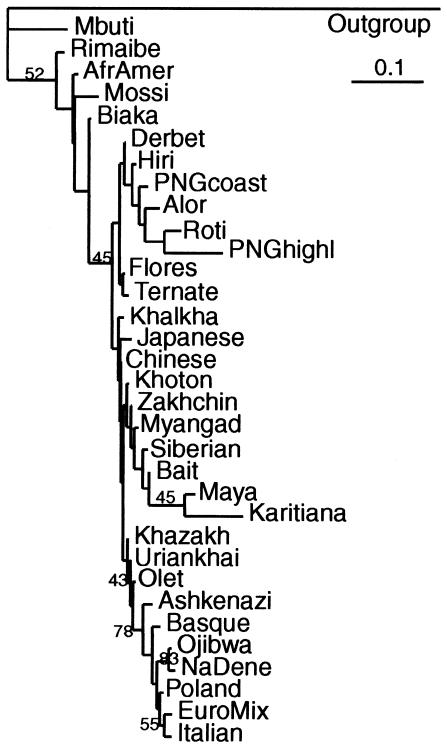
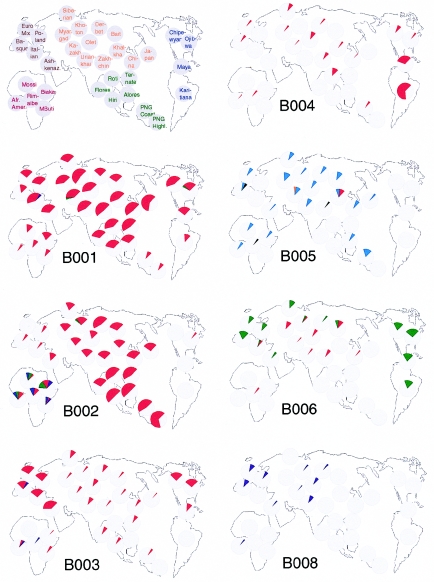
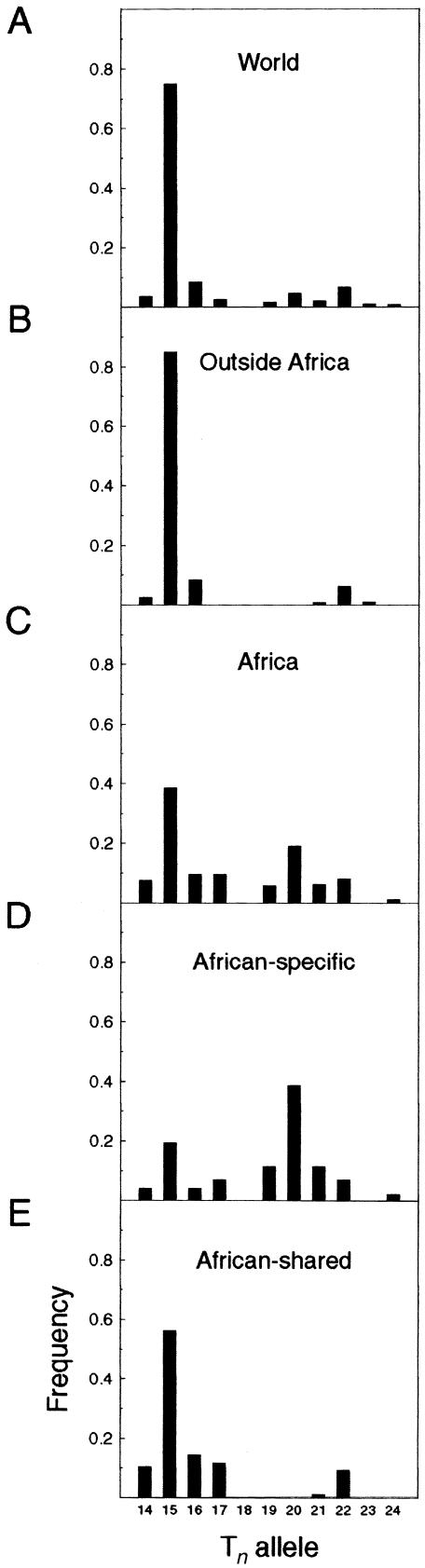
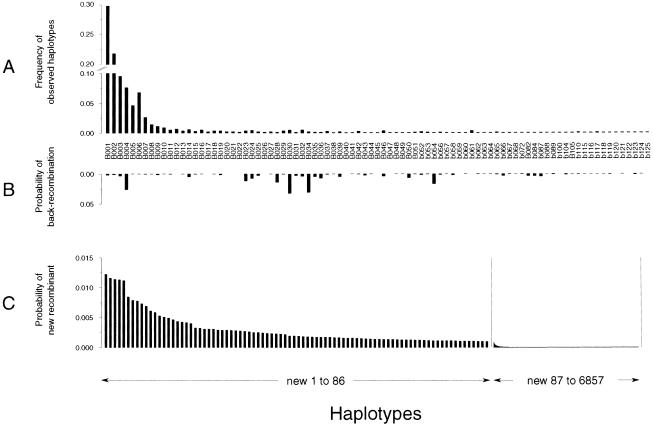
Although Africa has played a central role in human evolutionary history, certain studies have suggested that not all contemporary human genetic diversity is of recent African origin. We investigated 35 simple polymorphic sites and one T(n) microsatellite in an 8-kb segment of the dystrophin gene. We found 86 haplotypes in 1,343 chromosomes from around the world. Although a classical out-of-Africa topology was observed in trees based on the variant frequencies, the tree of haplotype sequences reveals three lineages accounting for present-day diversity. The proportion of new recombinants and the diversity of the T(n) microsatellite were used to estimate the age of haplotype lineages and the time of colonization events. The lineage that underwent the great expansion originated in Africa prior to the Upper Paleolithic (27,000-56,000 years ago). A second group, of structurally distinct haplotypes that occupy a central position on the tree, has never left Africa. The third lineage is represented by the haplotype that lies closest to the root, is virtually absent in Africa, and appears older than the recent out-of-Africa expansion. We propose that this lineage could have left Africa before the expansion (as early as 160,000 years ago) and admixed, outside of Africa, with the expanding lineage. Contemporary human diversity, although dominated by the recently expanded African lineage, thus represents a mosaic of different contributions.
Figures
References
Electronic-Database Information
-
- GenBank, http://www.ncbi.nlm.nih.gov/Genbank/ (for the human dystrophin gene [accession number U94396])
References
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
