

AMPA receptors mediate acetylcholine release from starburst amacrine cells in the rabbit retina
- PMID: 14515241
- PMCID: PMC3341736
- DOI: 10.1002/cne.10880
AMPA receptors mediate acetylcholine release from starburst amacrine cells in the rabbit retina
Abstract
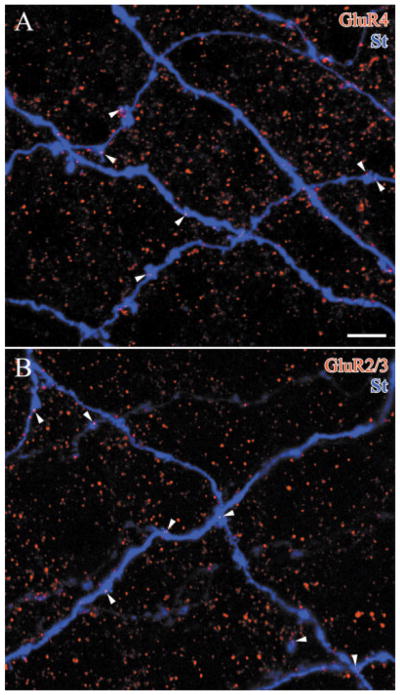
The light response of starburst amacrine cells is initiated by glutamate released from bipolar cells. To identify the receptors that mediate this response, we used a combination of anatomical and physiological techniques. An in vivo, rabbit eyecup was preloaded with [(3)H]-choline, and the [(3)H]-acetylcholine (ACh) released into the superfusate was monitored. A photopic, 3 Hz flashing light increased ACh release, and the selective AMPA receptor antagonist, GYKI 53655, blocked this light-evoked response. Nonselective AMPA/kainate agonists increased the release of ACh, but the specific kainate receptor agonist, SYM 2081, did not increase ACh release. Selective AMPA receptor antagonists, GYKI 53655 or GYKI 52466, also blocked the responses to agonists. We conclude that the predominant excitatory input to starburst amacrine cells is mediated by AMPA receptors. We also labeled lightly fixed rabbit retinas with antisera to choline acetyltransferase (ChAT), AMPA receptor subunits GluR1, GluR2/3, or GluR4, and kainate receptor subunits GluR6/7 and KA2. Labeled puncta were observed in the inner plexiform layer with each of these antisera to glutamate receptors, but only GluR2/3-IR puncta and GluR4-IR puncta were found on the ChAT-IR processes. The same was true of starburst cells injected intracellularly with Neurobiotin, and these AMPA receptor subunits were localized to two populations of puncta. The AMPA receptors are expected to desensitize rapidly, enhancing the sensitivity of starburst amacrine cells to moving or other rapidly changing stimuli.
Copyright 2003 Wiley-Liss, Inc.
Figures






References
-
- Ames A, Nesbett FB. In vitro retina as an experimental model of the central nervous system. J Neurochem. 1981;37:867–877. - PubMed
-
- Bleakman D, Ballyk BA, Schoepp DD, Palmer AJ, Bath CP, Sharpe EF, Woolley ML, Bufton HR, Kamboj RK, Tarnawa I, et al. Activity of 2,3-benzodiezepines at native rat and recombinant human glutamate receptors in vitro: stereospecificity and selectivity profiles. Neuropharmacology. 1996;35:1689–1702. - PubMed
-
- Brandon C. Cholinergic neurons in the rabbit retina: dendritic branching and ultrastructural connectivity. Brain Res. 1987;426:119–130. - PubMed
-
- Brown SP, Masland RH. Costratification of a population of bipolar cells with direction-selective circuitry of the rabbit retina. J Comp Neurol. 1999;408:97–106. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
