

High-resolution structure of the E.coli RecQ helicase catalytic core
- PMID: 14517231
- PMCID: PMC204483
- DOI: 10.1093/emboj/cdg500
High-resolution structure of the E.coli RecQ helicase catalytic core
Abstract
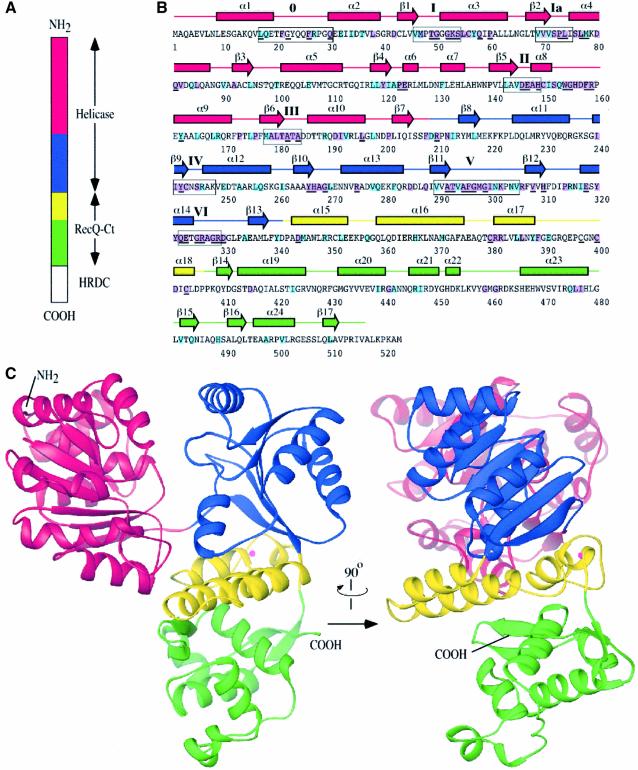
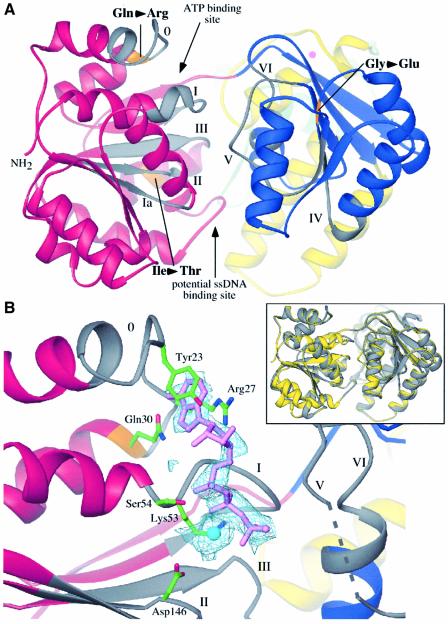
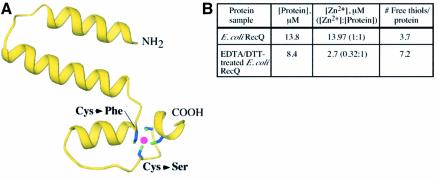
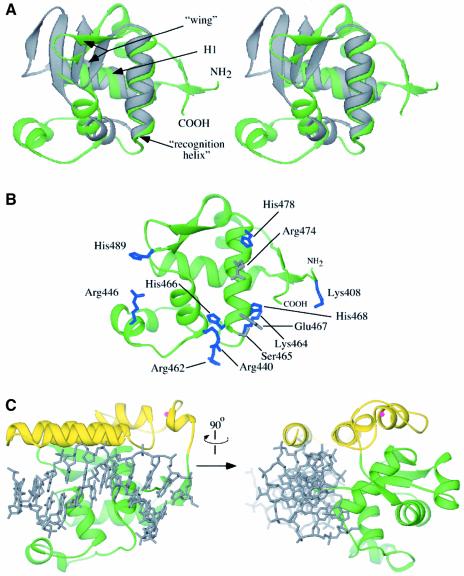
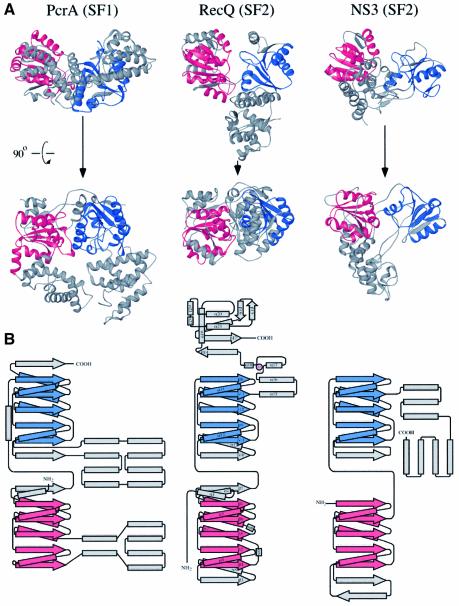
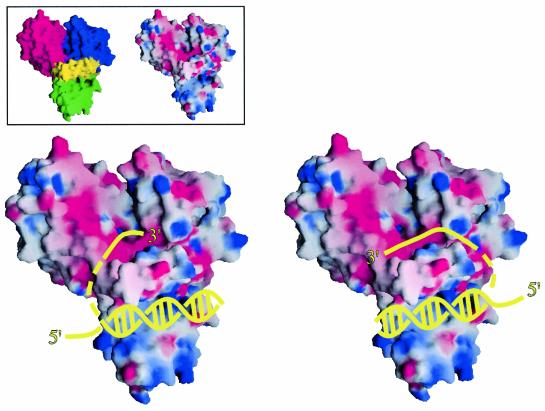
RecQ family helicases catalyze critical genome maintenance reactions in bacterial and eukaryotic cells, playing key roles in several DNA metabolic processes. Mutations in recQ genes are linked to genome instability and human disease. To define the physical basis of RecQ enzyme function, we have determined a 1.8 A resolution crystal structure of the catalytic core of Escherichia coli RecQ in its unbound form and a 2.5 A resolution structure of the core bound to the ATP analog ATPgammaS. The RecQ core comprises four conserved subdomains; two of these combine to form its helicase region, while the others form unexpected Zn(2+)-binding and winged-helix motifs. The structures reveal the molecular basis of missense mutations that cause Bloom's syndrome, a human RecQ-associated disease. Finally, based on findings from the structures, we propose a mechanism for RecQ activity that could explain its functional coordination with topoisomerase III.
Figures
References
-
- Bahr A., De Graeve,F., Kedinger,C. and Chatton,B. (1998) Point mutations causing Bloom’s syndrome abolish ATPase and DNA helicase activities of the BLM protein. Oncogene, 17, 2565–2571. - PubMed
-
- Barakat A., Ababou,M., Onclercq,R., Dutertre,S., Chadli,E., Hda,N., Benslimane,A. and Amor-Gueret,M. (2000) Identification of a novel BLM missense mutation (2706T>C) in a Moroccan patient with Bloom’s syndrome. Hum. Mutat., 15, 584–585. - PubMed
-
- Berg J.M. and Shi,Y. (1996) The galvanization of biology: a growing appreciation for the roles of zinc. Science, 271, 1081–1085. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
