

Regulation of V(D)J recombination by nucleosome positioning at recombination signal sequences
- PMID: 14517257
- PMCID: PMC204470
- DOI: 10.1093/emboj/cdg487
Regulation of V(D)J recombination by nucleosome positioning at recombination signal sequences
Abstract
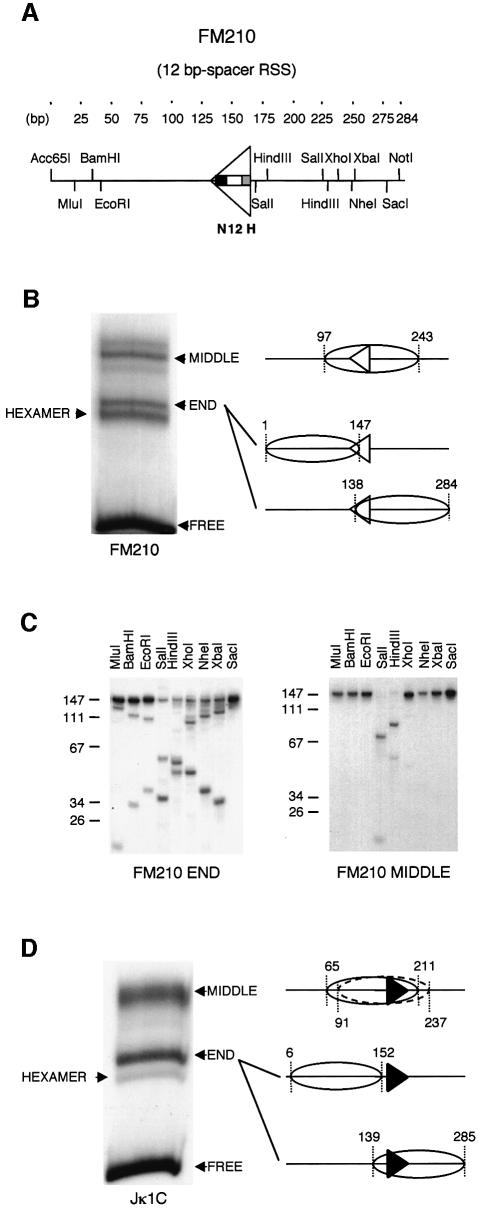
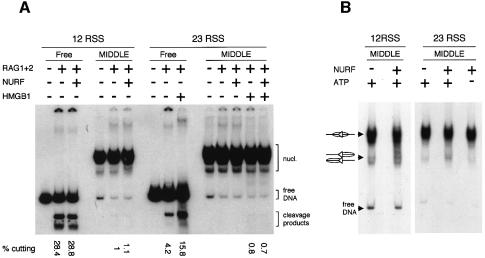
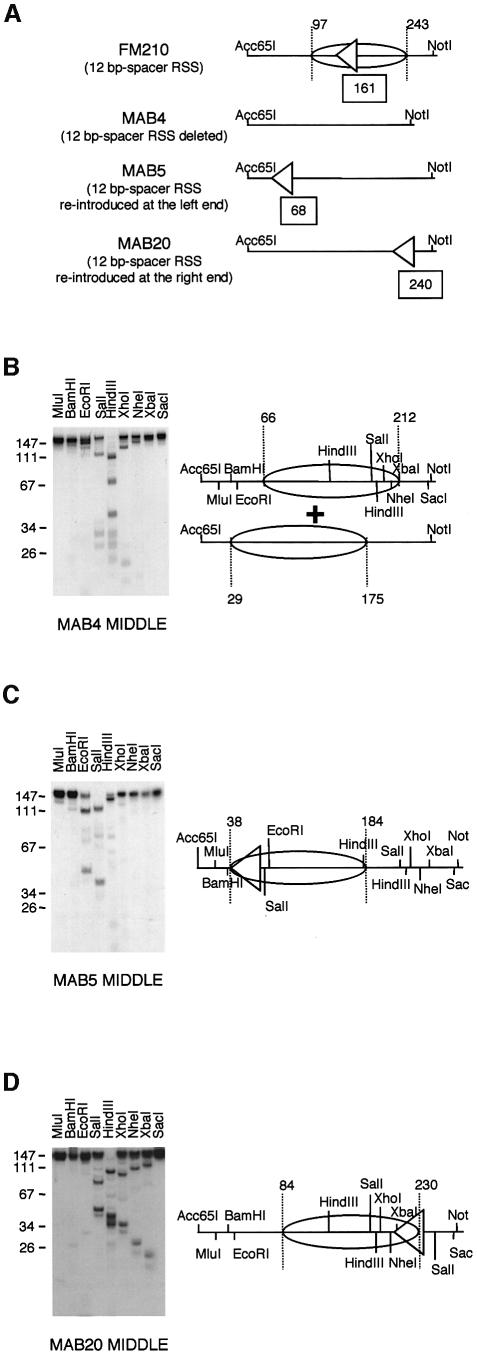
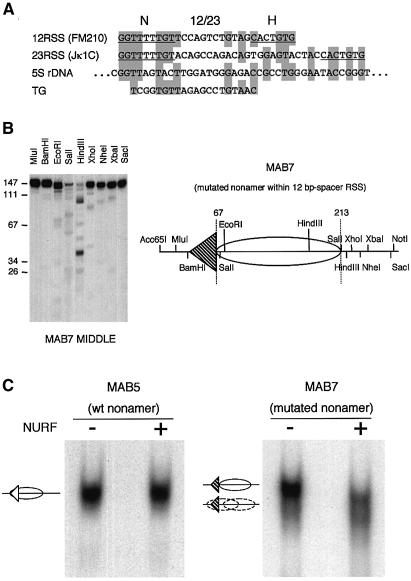
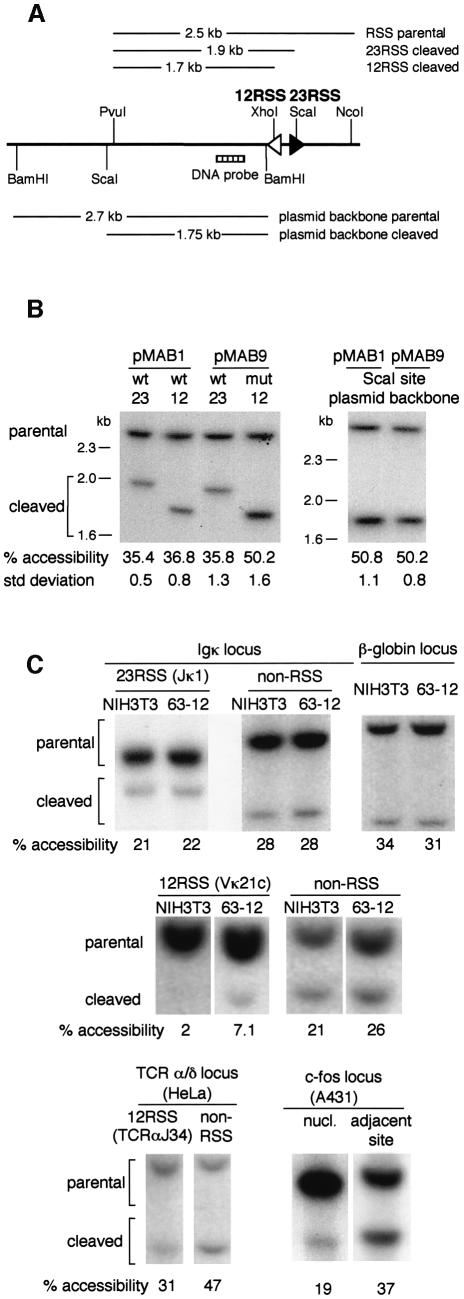
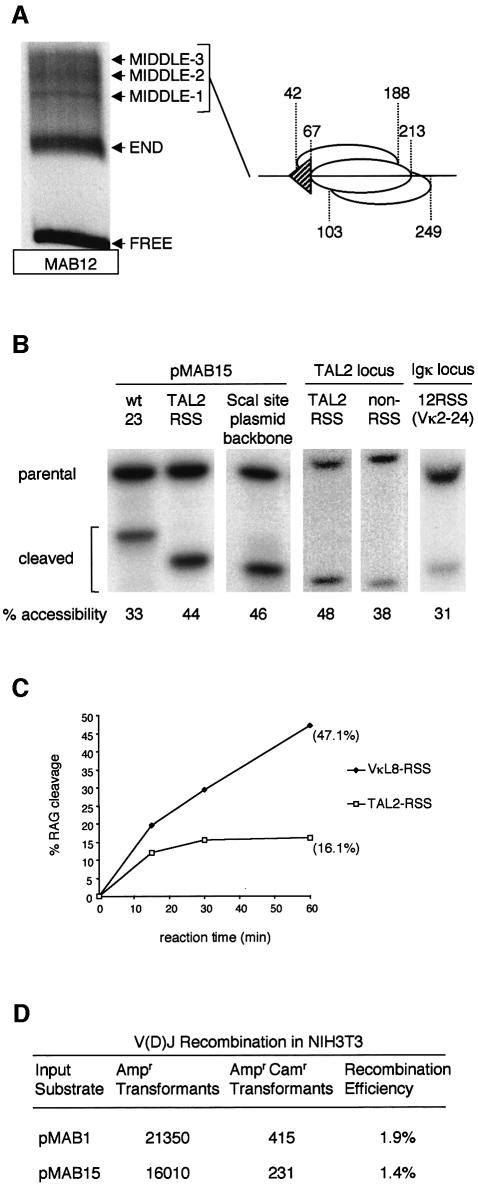
A key component in the regulation of V(D)J recombination is control of the accessibility of RAG proteins to recombination signal sequences (RSS). Nucleosomes are known to inhibit this accessibility. We show here that the signal sequence itself represses accessibility by causing nucleosome positioning over the RSS. This positioning is mediated, in vitro and in vivo, by the conserved nonamer of the RSS. Consistent with this strong positioning, nucleosomes at RSSs are resistant to remodelling by nucleosome sliding. In vivo we find that consensus RSSs are preferentially protected, whereas those that lack a consensus nonamer, including some cryptic RSSs, fail to position nucleosomes. Decreased protection of these non-consensus RSSs correlates with their increased use in recombination assays. We therefore suggest that nucleosome positioning by RSSs provides a previously unanticipated level of protection and regulation of V(D)J recombination.
Figures
References
-
- Bassing C.H., Swat,W. and Alt,F.W. (2002) The mechanism and regulation of chromosomal V(D)J recombination. Cell, 109 Suppl., S45–S55. - PubMed
-
- Boyes J. (1999) Preparation of chromatin templates for transcription studies. In Latchman,D.S. (ed.), Transcription Factors: A Practical Approach. Oxford University Press, Oxford, UK, pp. 229–260.
-
- Cairns B.R. and Kingston,R.E. (2000) The SWI/SNF family of remodelling complexes. In Elgin,S.C.R. and Workman,J.L. (eds), Chromatin Structure and Gene Expression. Oxford University Press, New York, NY, pp. 97–113.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
