

Unfolding pathways of native bacteriorhodopsin depend on temperature
- PMID: 14517259
- PMCID: PMC204492
- DOI: 10.1093/emboj/cdg509
Unfolding pathways of native bacteriorhodopsin depend on temperature
Abstract
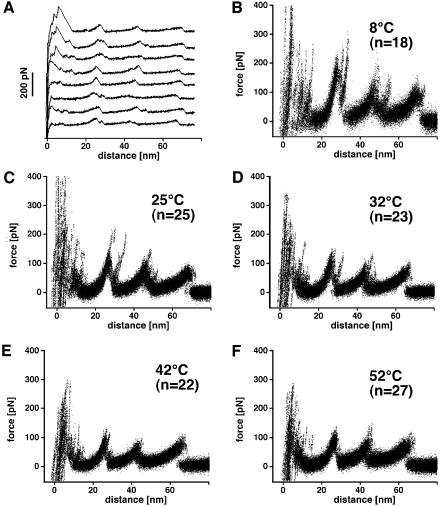
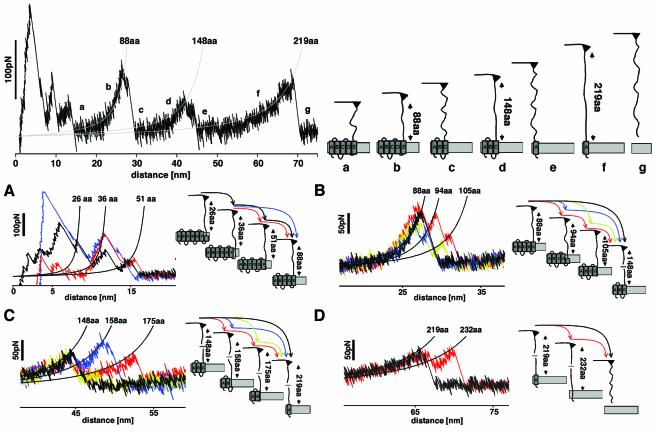
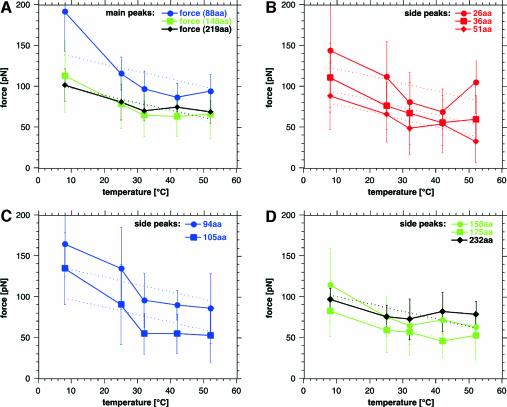
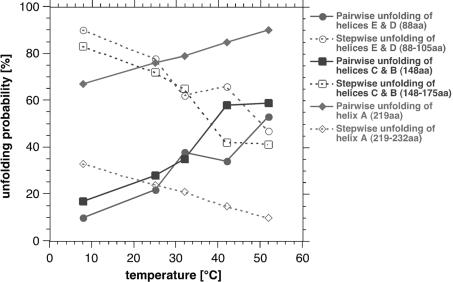
The combination of high-resolution atomic force microscopy (AFM) imaging and single-molecule force-spectroscopy was employed to unfold single bacteriorhodopsins (BR) from native purple membrane patches at various physiologically relevant temperatures. The unfolding spectra reveal detailed insight into the stability of individual structural elements of BR against mechanical unfolding. Intermittent states in the unfolding process are associated with the stepwise unfolding of alpha-helices, whereas other states are associated with the unfolding of polypeptide loops connecting the alpha-helices. It was found that the unfolding forces of the secondary structures considerably decreased upon increasing the temperature from 8 to 52 degrees C. Associated with this effect, the probability of individual unfolding pathways of BR was significantly influenced by the temperature. At lower temperatures, transmembrane alpha-helices and extracellular polypeptide loops exhibited sufficient stability to individually establish potential barriers against unfolding, whereas they predominantly unfolded collectively at elevated temperatures. This suggests that increasing the temperature decreases the mechanical stability of secondary structural elements and changes molecular interactions between secondary structures, thereby forcing them to act as grouped structures.
Figures
References
-
- Bell G.I. (1978) Models for the specific adhesion of cells to cells. Science, 200, 618–627. - PubMed
-
- Belrhali H., Nollert,P., Royant,A., Menzel,C., Rosenbusch,J.P., Landau,E.M. and Pebay-Peyroula,E. (1999) Protein, lipid and water organization in bacteriorhodopsin crystals: a molecular view of the purple membrane at 1.9 Å resolution. Structure Fold. Des., 7, 909–917. - PubMed
-
- Blume A. (1983) Apparent molar heat capacities of phospholipids in aqueous dispersion: effects of chain length and head group structure. Biochemistry, 22, 5436–5442.
-
- Booth P.J., Templer,R.H., Meijberg,W., Allen,S.J., Curran,A.R. and Lorch,M. (2001) In vitro studies of membrane protein folding. Crit. Rev. Biochem. Mol. Biol., 36, 501–603. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
