

Daxx silencing sensitizes cells to multiple apoptotic pathways
- PMID: 14517282
- PMCID: PMC230316
- DOI: 10.1128/MCB.23.20.7108-7121.2003
Daxx silencing sensitizes cells to multiple apoptotic pathways
Abstract
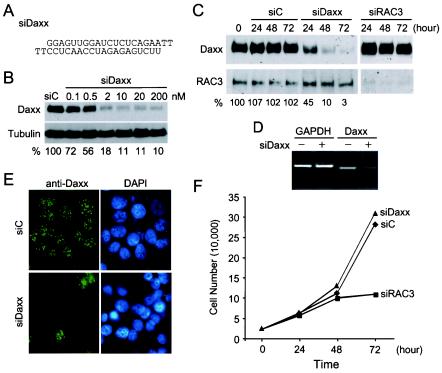
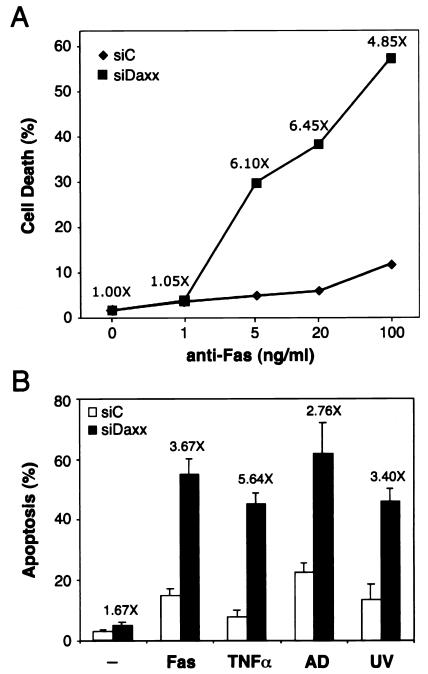
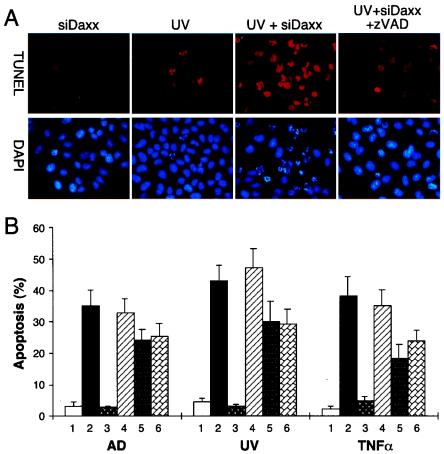
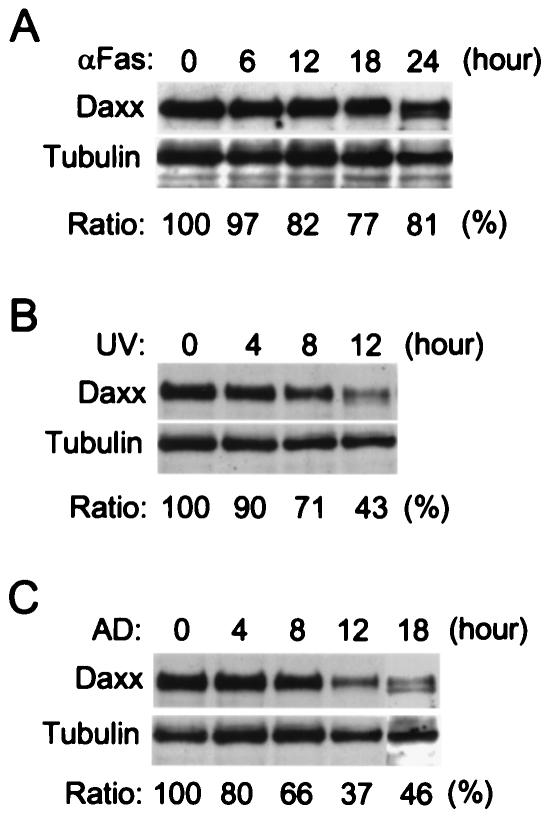
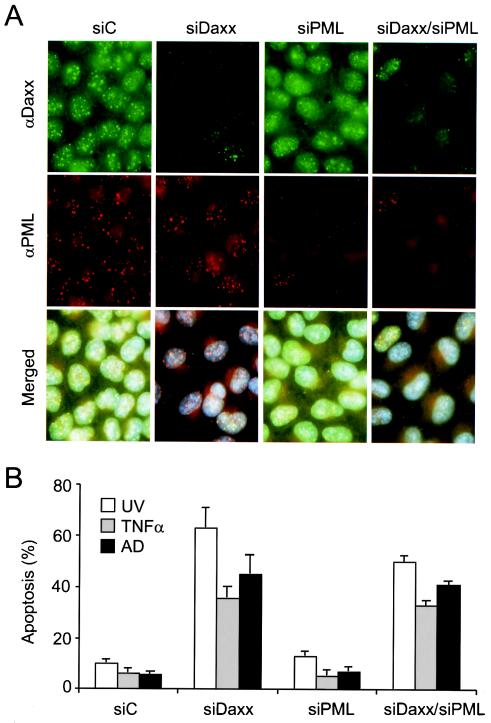
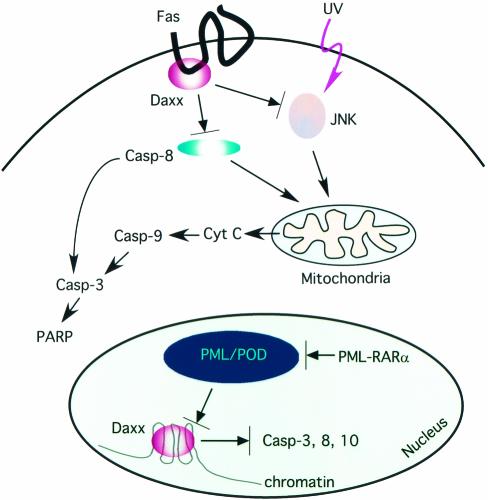
Daxx is a nuclear protein involved in apoptosis and transcriptional repression, and it interacts with the death receptor Fas, promyelocytic leukemia protein (PML), and several transcriptional repressors. The function of Daxx in apoptosis is controversial because opposite results were obtained in transient overexpression and genetic knockout studies. Furthermore, the roles of PML and transcriptional repression in Daxx-regulated apoptosis are currently unknown. In this study, we investigated the role of Daxx in Fas- and stress-induced apoptosis by small interfering RNA-mediated Daxx silencing in mammalian cells. Daxx silencing had no apparent cytotoxic effects on mammalian cells within 72 h. Intriguingly, Daxx silencing strongly sensitized cells to Fas- and stress-induced apoptosis, which was accompanied by caspase activation, cytochrome c release, and Jun N-terminal kinase activation. Consistently, endogenous Daxx was degraded rapidly upon induction of apoptosis by stress or anti-Fas antibody. Finally, PML silencing had no effect on Daxx silencing-mediated apoptotic events, while caspase gene expression was upregulated in the absence of Daxx. These data strongly suggest that Daxx may inhibit Fas and stress-mediated apoptosis by suppressing proapoptotic gene expression outside of PML domains.
Figures




References
-
- Amin, H. M., S. Saeed, and S. Alkan. 2001. Histone deacetylase inhibitors induce caspase-dependent apoptosis and downregulation of daxx in acute promyelocytic leukaemia with t(15;17). Br. J. Haematol. 115:287-297. - PubMed
-
- Cermak, L., a. imova, A. Pintzas, V. Horeji, and L. Andera. 2002. Molecular mechanisms involved in CD43-mediated apoptosis of TF-1 cells: roles of transcription, Daxx expression and adhesion molecules. J. Biol. Chem. 277:7955-7961. - PubMed
-
- Chang, H. Y., H. Nishitoh, X. Yang, H. Ichijo, and D. Baltimore. 1998. Activation of apoptosis signal- regulating kinase 1 (ASK1) by the adapter protein daxx. Science 281:1860-1863. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous