

Time- and exercise-dependent gene regulation in human skeletal muscle
- PMID: 14519196
- PMCID: PMC328450
- DOI: 10.1186/gb-2003-4-10-r61
Time- and exercise-dependent gene regulation in human skeletal muscle
Abstract
Background: Skeletal muscle remodeling is a critical component of an organism's response to environmental changes. Exercise causes structural changes in muscle and can induce phase shifts in circadian rhythms, fluctuations in physiology and behavior with a period of around 24 hours that are maintained by a core clock mechanism. Both exercise-induced remodeling and circadian rhythms rely on the transcriptional regulation of key genes.
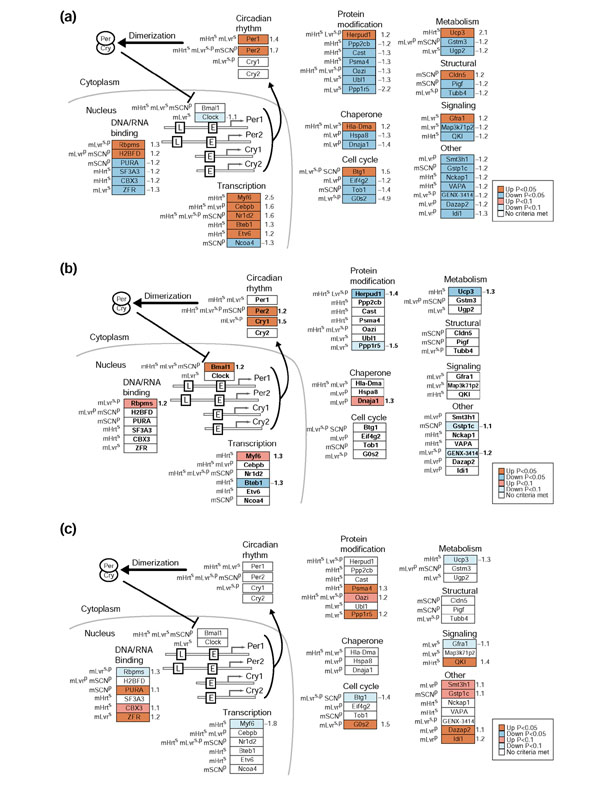
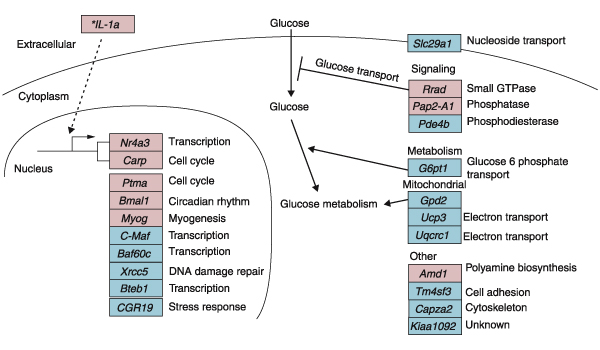
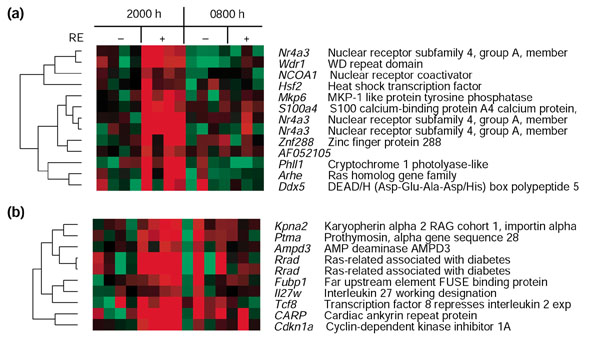
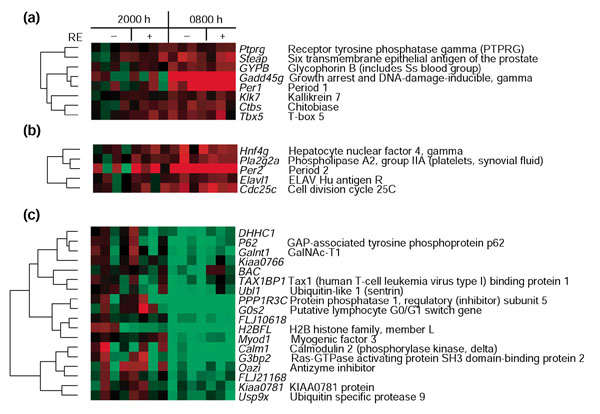
Results: We used DNA microarrays to determine the effects of resistance exercise (RE) on gene regulation in biopsy samples of human quadriceps muscle obtained 6 and 18 hours after an acute bout of isotonic exercise with one leg. We also profiled diurnal gene regulation at the same time points (2000 and 0800 hours) in the non-exercised leg. Comparison of our results with published circadian gene profiles in mice identified 44 putative genes that were regulated in a circadian fashion. We then used quantitative PCR to validate the circadian expression of selected gene orthologs in mouse skeletal muscle.
Conclusions: The coordinated regulation of the circadian clock genes Cry1, Per2, and Bmal1 6 hours after RE and diurnal genes 18 hours after RE in the exercised leg suggest that RE may directly modulate circadian rhythms in human skeletal muscle.
Figures



References
-
- Ryan AS. Insulin resistance with aging: effects of diet and exercise. Sports Med. 2000;30:327–346. - PubMed
-
- Pollock ML, Franklin BA, Balady GJ, Chaitman BL, Fleg JL, Fletcher B, Limacher M, Piña IL, Stein RA, Williams M, Bazzarre T. AHA Science Advisory. Resistance exercise in individuals with and without cardiovascular disease: benefits, rationale, safety, and prescription. An advisory from the Committee on Exercise, Rehabilitation, and Prevention, Council on Clinical Cardiology, American Heart Association; Position paper endorsed by the American College of Sports Medicine. Circulation. 2000;101:828–833. - PubMed
-
- Baar K, Esser K. Phosphorylation of p70(S6k) correlates with increased skeletal muscle mass following resistance exercise. Am J Physiol. 1999;276:C120–C127. - PubMed
-
- McCall GE, Byrnes WC, Dickinson A, Pattany PM, Fleck SJ. Muscle fiber hypertrophy, hyperplasia, and capillary density in college men after resistance training. J Appl Physiol. 1996;81:2004–2012. - PubMed
-
- Dusterhoft S, Putman CT, Pette D. Changes in FGF and FGF receptor expression in low-frequency-stimulated rat muscles and rat satellite cell cultures. Differentiation. 1999;65:203–208. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
