

Brassinosteroids promote root growth in Arabidopsis
- PMID: 14526105
- PMCID: PMC281621
- DOI: 10.1104/pp.103.028662
Brassinosteroids promote root growth in Arabidopsis
Abstract
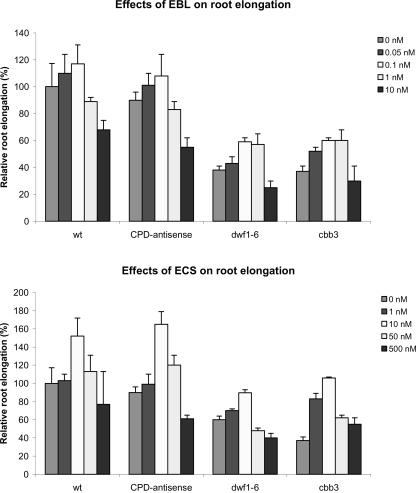
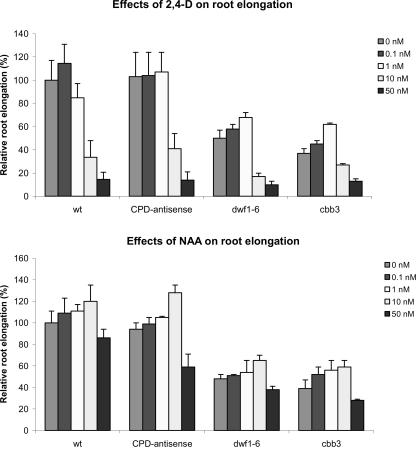
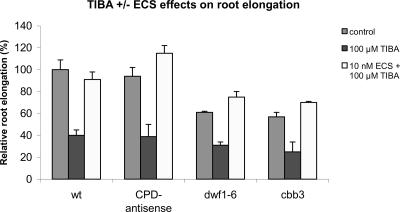
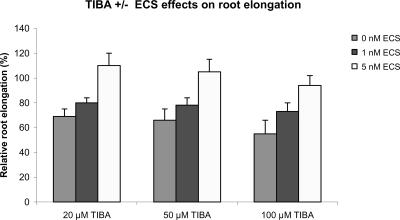
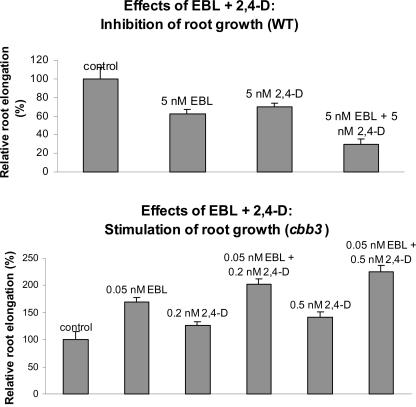
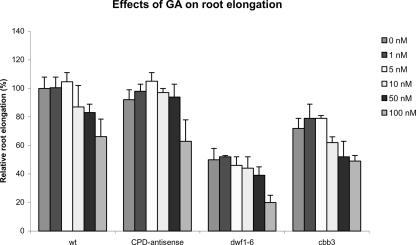
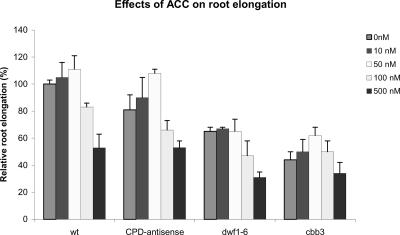
Although brassinosteroids (BRs) are known to regulate shoot growth, their role in the regulation of root growth is less clear. We show that low concentrations of BRs such as 24-epicastasterone and 24-epibrassinolide promote root elongation in Arabidopsis wild-type plants up to 50% and in BR-deficient mutants such as dwf1-6 (cbb1) and cbb3 (which is allelic to cpd) up to 150%. The growth-stimulating effect of exogenous BRs is not reduced by the auxin transport inhibitor 2,3,5-triidobenzoic acid. BR-deficient mutants show normal gravitropism, and 2,3,5-triidobenzoic acid or higher concentrations of 2,4-dichlorophenoxyacetic acid and naphtaleneacetic acid inhibit root growth in the mutants to the same extent as in wild-type plants. Simultaneous administration of 24-epibrassinolide and 2,4-dichlorophenoxyacetic acid results in largely additive effects. Exogenous gibberellins do not promote root elongation in the BR-deficient mutants, and the sensitivity to the ethylene precursor 1-aminocyclopropane-1-carboxylic acid is not altered. Thus, the root growth-stimulating effect of BRs appears to be largely independent of auxin and gibberellin action. Furthermore, we analyzed BR interactions with other phytohormones on the gene expression level. Only a limited set of auxin- and ethylene-related genes showed altered expression levels. Genes related to other phytohormones barely showed changes, providing further evidence for an autonomous stimulatory effect of BR on root growth.
Figures
Similar articles
-
Brassinosteroids interact with auxin to promote lateral root development in Arabidopsis.Plant Physiol. 2004 Apr;134(4):1624-31. doi: 10.1104/pp.103.036897. Epub 2004 Mar 26. Plant Physiol. 2004. PMID: 15047895 Free PMC article.
-
Arabidopsis constitutive photomorphogenic mutant, bls1, displays altered brassinosteroid response and sugar sensitivity.Plant Mol Biol. 2004 Sep;56(2):185-201. doi: 10.1007/s11103-004-2799-x. Plant Mol Biol. 2004. PMID: 15604737
-
Brassinosteroids stimulate plant tropisms through modulation of polar auxin transport in Brassica and Arabidopsis.Plant Cell. 2005 Oct;17(10):2738-53. doi: 10.1105/tpc.105.034397. Epub 2005 Sep 2. Plant Cell. 2005. PMID: 16141452 Free PMC article.
-
Brassinosteroids and plant steroid hormone signaling.Plant Cell. 2002;14 Suppl(Suppl):S97-110. doi: 10.1105/tpc.001461. Plant Cell. 2002. PMID: 12045272 Free PMC article. Review. No abstract available.
-
Biological effects of brassinosteroids.Crit Rev Biochem Mol Biol. 1999;34(5):339-58. doi: 10.1080/10409239991209345. Crit Rev Biochem Mol Biol. 1999. PMID: 10565679 Review. No abstract available.
Cited by
-
Spatiotemporal brassinosteroid signaling and antagonism with auxin pattern stem cell dynamics in Arabidopsis roots.Curr Biol. 2015 Apr 20;25(8):1031-42. doi: 10.1016/j.cub.2015.02.046. Epub 2015 Apr 9. Curr Biol. 2015. PMID: 25866388 Free PMC article.
-
Ethylene and Hormonal Cross Talk in Vegetative Growth and Development.Plant Physiol. 2015 Sep;169(1):61-72. doi: 10.1104/pp.15.00724. Epub 2015 Jul 31. Plant Physiol. 2015. PMID: 26232489 Free PMC article. Review.
-
The Research Process of PSK Biosynthesis, Signaling Transduction, and Potential Applications in Brassica napus.Plants (Basel). 2023 Aug 28;12(17):3075. doi: 10.3390/plants12173075. Plants (Basel). 2023. PMID: 37687322 Free PMC article. Review.
-
Quantification of the brassinosteroid insensitive1 receptor in planta.Plant Physiol. 2011 Aug;156(4):1691-700. doi: 10.1104/pp.111.179309. Epub 2011 May 26. Plant Physiol. 2011. PMID: 21617031 Free PMC article.
-
Overexpression of DWF1 Enhances Low-Nitrogen Stress Tolerance in Potato Plants.Int J Mol Sci. 2025 May 4;26(9):4374. doi: 10.3390/ijms26094374. Int J Mol Sci. 2025. PMID: 40362610 Free PMC article.
References
-
- Arteca JM, Arteca RN (2001) Brassinosteroid-induced exaggerated growth in hydroponically grown Arabidopsis plants. Physiol Plant 112: 104–112 - PubMed
-
- Arteca RN, Bachman JM, Yopp JH, Mandava NB (1985) Relationship of steroidal structure to ethylene production by etiolated mung bean segments. Physiol Plant 64: 13–16
-
- Bertell G, Eliasson L (1992) Cytokinin effects on root growth and possible interactions with ethylene and indole-3-acetic acid. Physiol Plant 84: 255–261
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
