

Membrane association of myotubularin-related protein 2 is mediated by a pleckstrin homology-GRAM domain and a coiled-coil dimerization module
- PMID: 14530412
- PMCID: PMC218732
- DOI: 10.1073/pnas.2132732100
Membrane association of myotubularin-related protein 2 is mediated by a pleckstrin homology-GRAM domain and a coiled-coil dimerization module
Abstract
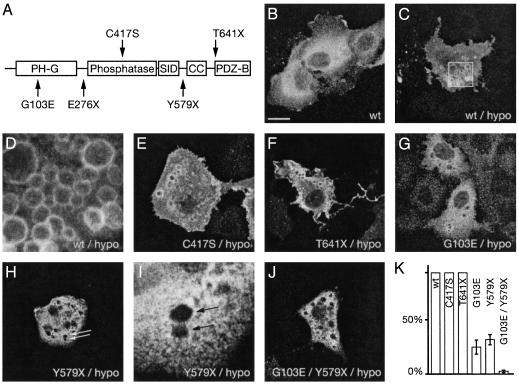
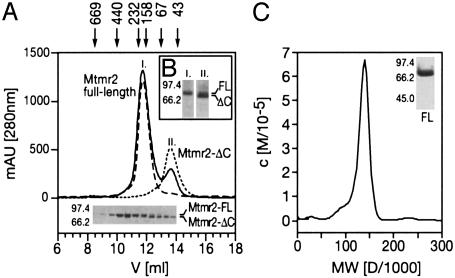
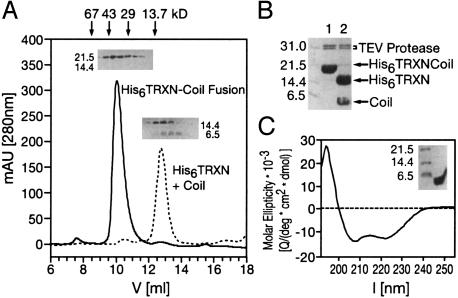
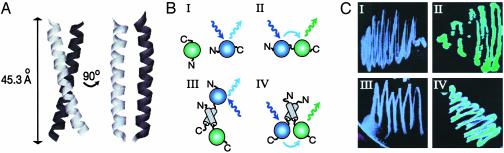
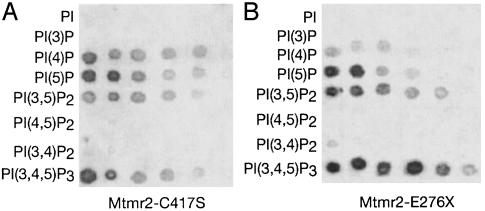
Mutations in the myotubularin (MTM)-related protein 2 (MTMR2) gene are responsible for the severe autosomal recessive neuropathy Charcot-Marie-Tooth disease type 4B1. MTMR2 belongs to the MTM family of dual-specific phosphatases that use phosphatidylinositol (PI) 3,5-bisphosphate [PI(3,5)P2] and PI 3-phosphate [PI(3)P] as their substrate. Because these substrates are localized in the membrane bilayer, membrane targeting of Mtmr2 is an important regulatory mechanism. In hypoosmotically stressed COS cells with increased levels of PI(3,5)P2, Mtmr2 is bound to the membrane of vacuoles formed under these conditions. Using several mutant forms of Mtmr2, we identified two domains that are necessary for membrane association: (i) A pleckstrin homology-GRAM domain; and (ii) a coiled-coil module. Protein-lipid overlay assays show that the pleckstrin homology-GRAM domain binds to PI(3,5)P2 and PI(5)P, a substrate and a product of the Mtmr2 enzyme, respectively. We also demonstrate that Mtmr2 forms a dimer and that the C-terminal coiled-coil is responsible for homodimerization, in addition to membrane association. Our data indicate that phosphoinositide-protein interactions, as well as protein-protein interactions, are necessary for the correct regulation of MTMR2.
Figures
References
-
- Wishart, M. J. & Dixon, J. E. (2002) Trends Cell Biol. 12, 579–585. - PubMed
-
- Bolino, A., Muglia, M., Conforti, F. L., LeGuern, E., Salih, M. A., Georgiou, D. M., Christodoulou, K., Hausmanowa-Petrusewicz, I., Mandich, P., Schenone, A., et al. (2000) Nat. Genet. 25, 17–19. - PubMed
-
- Houlden, H., King, R. H., Wood, N. W., Thomas, P. K. & Reilly, M. M. (2001) Brain 124, 907–915. - PubMed
-
- Bolino, A., Lonie, L. J., Zimmer, M., Boerkoel, C. F., Takashima, H., Monaco, A. P. & Lupski, J. R. (2001) Neurogenetics 3, 107–109. - PubMed
-
- Nelis, E., Erdem, S., Tan, E., Lofgren, A., Ceuterick, C., De Jonghe, P., Van Broeckhoven, C., Timmerman, V. & Topaloglu, H. (2002) Neuromuscular Disord. 12, 869–873. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
