

Loss of K-Cl co-transporter KCC3 causes deafness, neurodegeneration and reduced seizure threshold
- PMID: 14532115
- PMCID: PMC213773
- DOI: 10.1093/emboj/cdg519
Loss of K-Cl co-transporter KCC3 causes deafness, neurodegeneration and reduced seizure threshold
Abstract
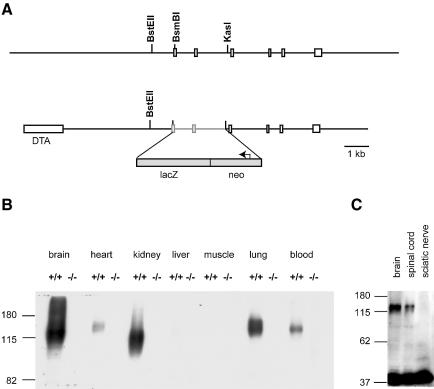
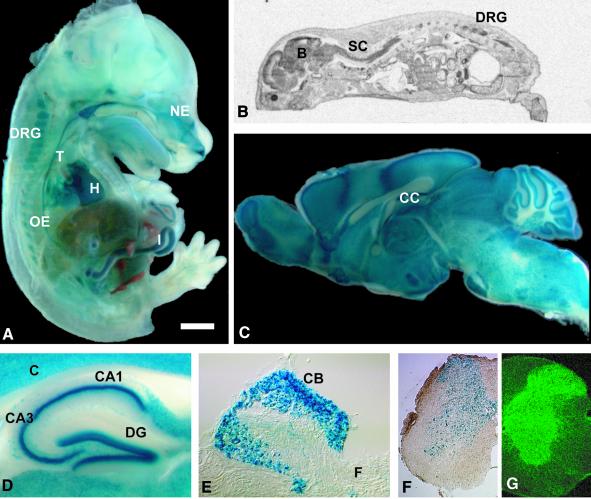
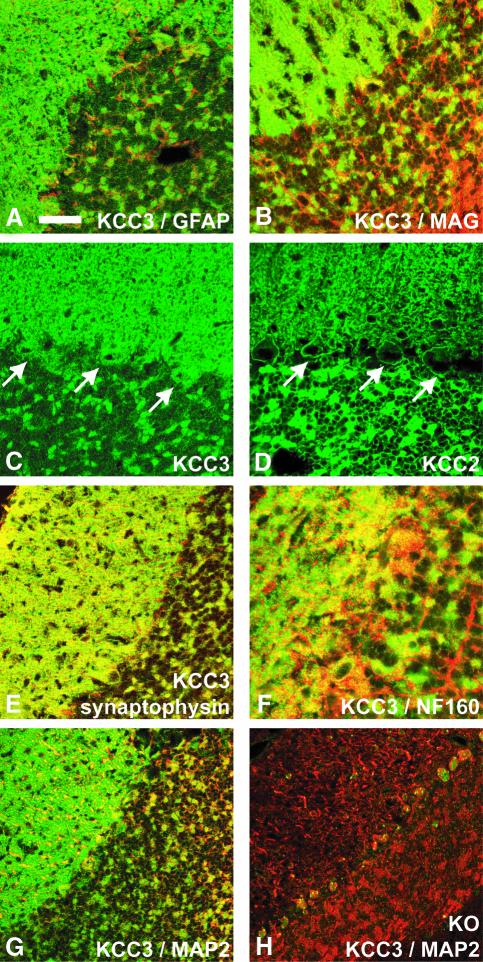
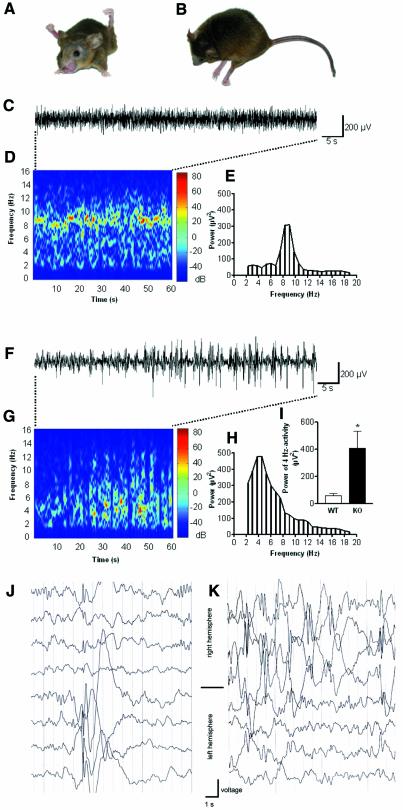
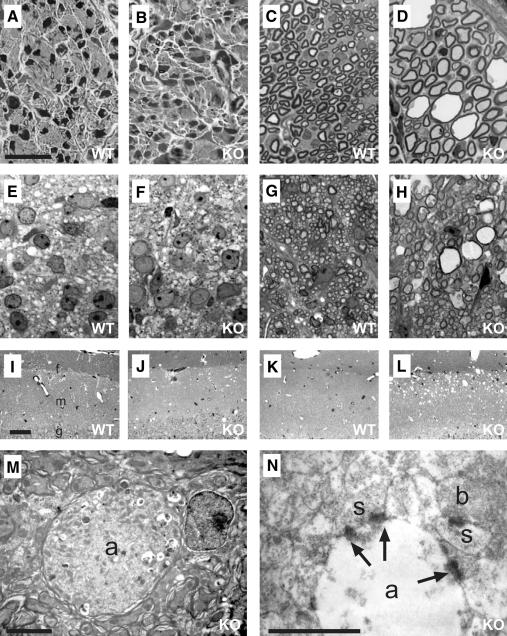
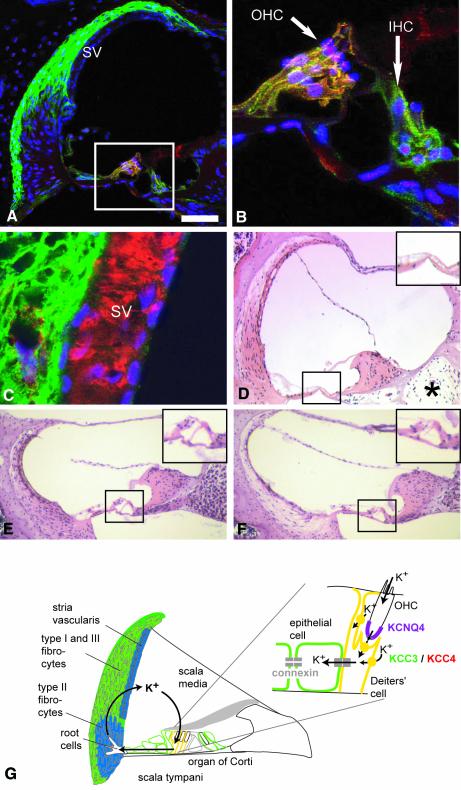
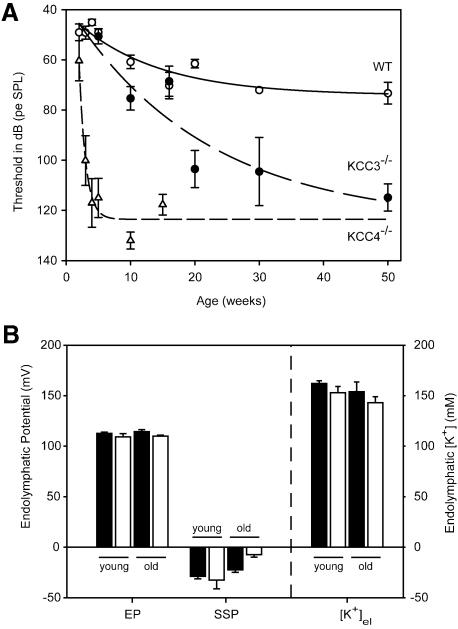
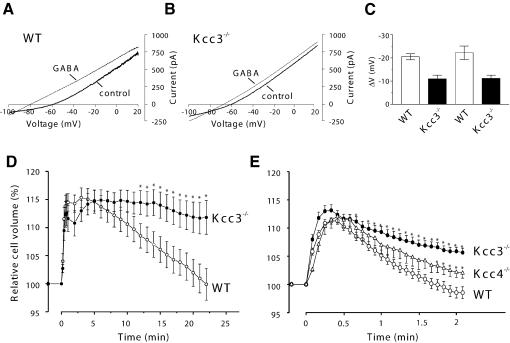
K-Cl co-transporters are encoded by four homologous genes and may have roles in transepithelial transport and in the regulation of cell volume and cytoplasmic chloride. KCC3, an isoform mutated in the human Anderman syndrome, is expressed in brain, epithelia and other tissues. To investigate the physiological functions of KCC3, we disrupted its gene in mice. This severely impaired cell volume regulation as assessed in renal tubules and neurons, and moderately raised intraneuronal Cl(-) concentration. Kcc3(-/-) mice showed severe motor abnormalities correlating with a progressive neurodegeneration in the peripheral and CNS. Although no spontaneous seizures were observed, Kcc3(-/-) mice displayed reduced seizure threshold and spike-wave complexes on electrocorticograms. These resembled EEG abnormalities in patients with Anderman syndrome. Kcc3(-/-) mice also displayed arterial hypertension and a slowly progressive deafness. KCC3 was expressed in many, but not all cells of the inner ear K(+) recycling pathway. These cells slowly degenerated, as did sensory hair cells. The present mouse model has revealed important cellular and systemic functions of KCC3 and is highly relevant for Anderman syndrome.
Figures
References
-
- Ando M. and Takeuchi,S. (1999) Immunological identification of an inward rectifier K+ channel (Kir4.1) in the intermediate cell (melanocyte) of the cochlear stria vascularis of gerbils and rats. Cell Tissue Res., 298, 179–183. - PubMed
-
- Boettger T., Hübner,C.A., Maier,H., Rust,M.B., Beck,F.X. and Jentsch,T.J. (2002) Deafness and renal tubular acidosis in mice lacking the K-Cl co-transporter Kcc4. Nature, 416, 874–878. - PubMed
-
- Crunelli V. and Leresche,N. (2002) Childhood absence epilepsy: genes, channels, neurons and networks. Nat. Rev. Neurosci., 3, 371–382. - PubMed
-
- Dupré N., Howard,H.C., Mathieu,J., Karpati,G., Vanasse,M., Bouchard,J.P., Carpenter,S. and Rouleau,G.A. (2003) Hereditary motor and sensory neuropathy with agenesis of the corpus callosum. Ann. Neurol., 54, 9–18. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
