

CHIP activates HSF1 and confers protection against apoptosis and cellular stress
- PMID: 14532117
- PMCID: PMC213783
- DOI: 10.1093/emboj/cdg529
CHIP activates HSF1 and confers protection against apoptosis and cellular stress
Abstract
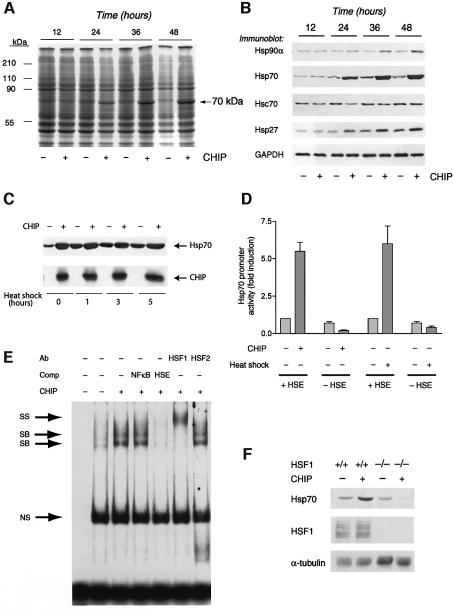
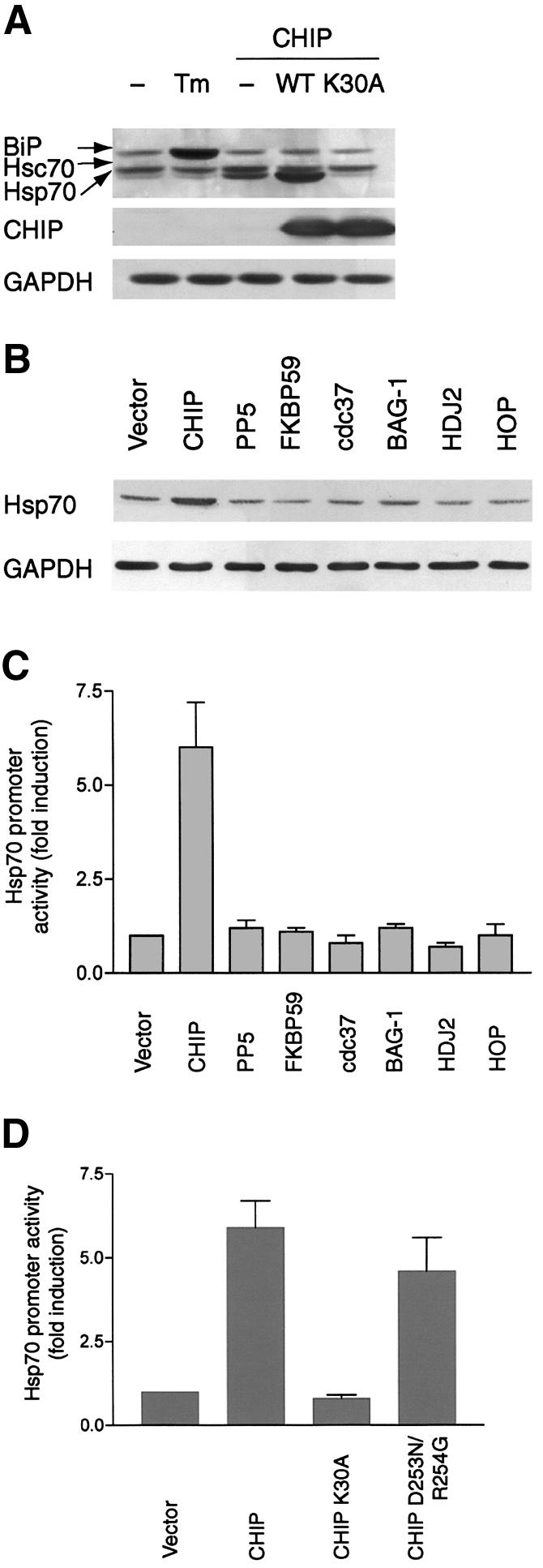
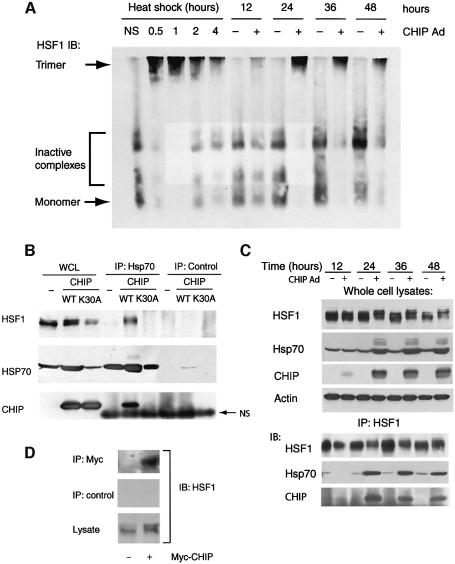
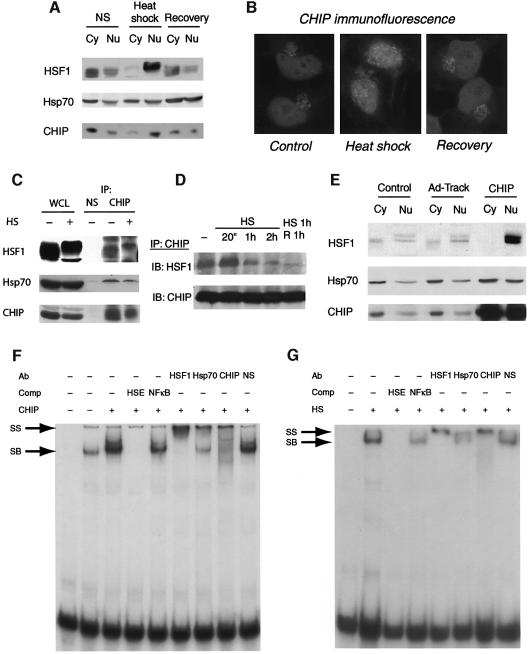
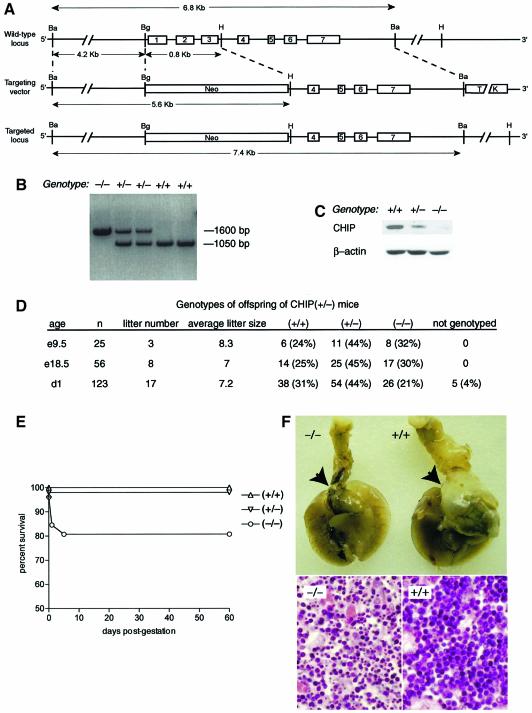
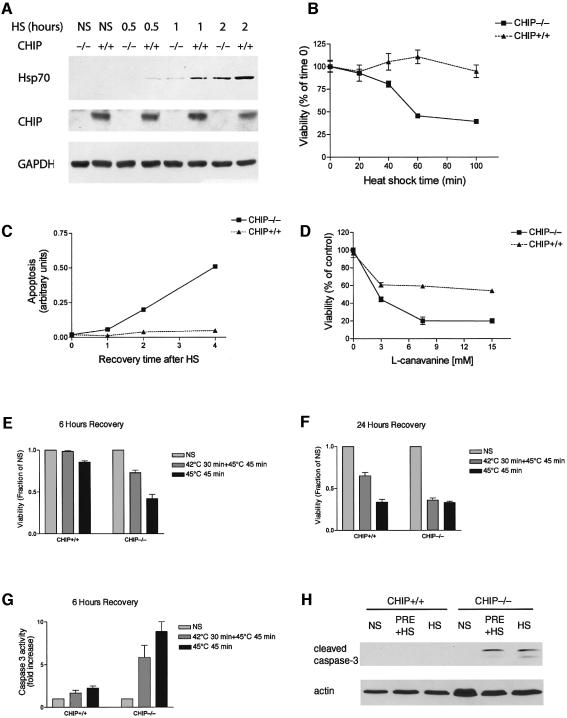
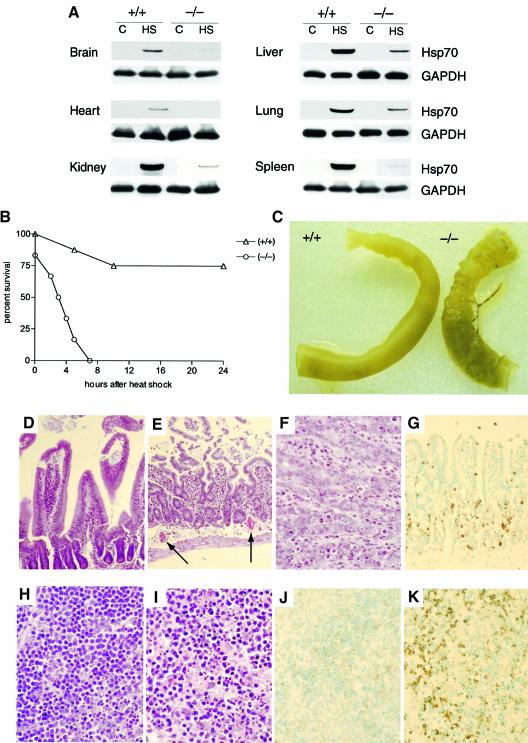
Induction of molecular chaperones is the characteristic protective response to environmental stress, and is regulated by a transcriptional program that depends on heat shock factor 1 (HSF1), which is normally under negative regulatory control by molecular chaperones Hsp70 and Hsp90. In metazoan species, the chaperone system also provides protection against apoptosis. We demonstrate that the dual function co-chaperone/ubiquitin ligase CHIP (C-terminus of Hsp70-interacting protein) regulates activation of the stress-chaperone response through induced trimerization and transcriptional activation of HSF1, and is required for protection against stress-induced apoptosis in murine fibroblasts. The consequences of this function are demonstrated by the phenotype of mice lacking CHIP, which develop normally but are temperature-sensitive and develop apoptosis in multiple organs after environmental challenge. CHIP exerts a central and unique role in tuning the response to stress at multiple levels by regulation of protein quality control and transcriptional activation of stress response signaling.
Figures
References
-
- Aravind L. and Koonin,E.V. (2000) The U box is a modified RING finger—a common domain in ubiquitination. Curr. Biol., 10, R132–R134. - PubMed
-
- Ballinger C.A., Connell,P., Wu,Y., Hu,Z., Thompson,L.J., Yin,L.-Y. and Patterson,C. (1999) Identification of CHIP, a novel tetratricopeptide repeat-containing protein that interacts with heat shock proteins and negatively regulates chaperone functions. Mol. Cell. Biol., 19, 4535–4545. - PMC - PubMed
-
- Beere H.M. and Green,D.R. (2001) Stress management: heat shock protein-70 and the regulation of apoptosis. Trends Cell Biol., 11, 6–10. - PubMed
-
- Bercovich B., Stancovski,I., Mayer,A., Blumenfeld,N., Laszlo,A., Schwartz,A.L. and Ciechanover,A. (1997) Ubiquitin-dependent degradation of certain protein substrates in vitro requires the molecular chaperone Hsc70. J. Biol. Chem., 272, 9002–9010. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
