

Impaired long-term memory and NR2A-type NMDA receptor-dependent synaptic plasticity in mice lacking c-Fos in the CNS
- PMID: 14534245
- PMCID: PMC6740829
- DOI: 10.1523/JNEUROSCI.23-27-09116.2003
Impaired long-term memory and NR2A-type NMDA receptor-dependent synaptic plasticity in mice lacking c-Fos in the CNS
Abstract
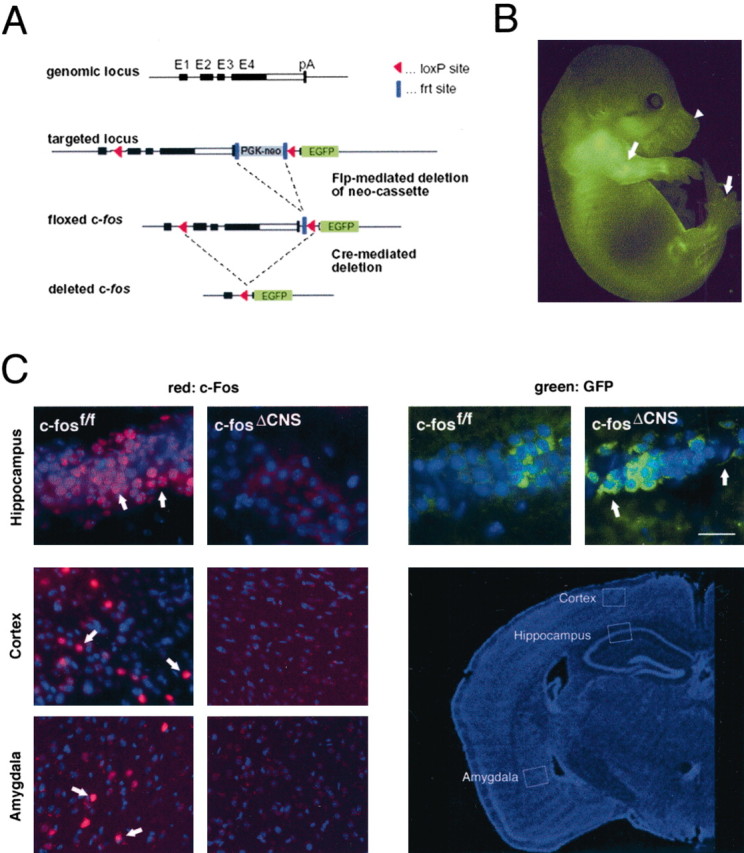
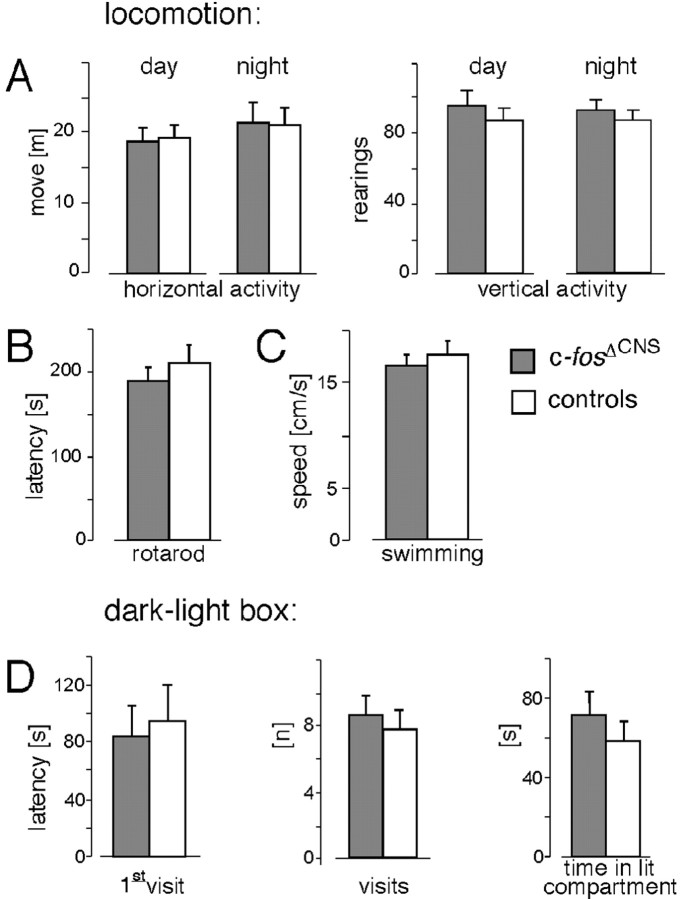
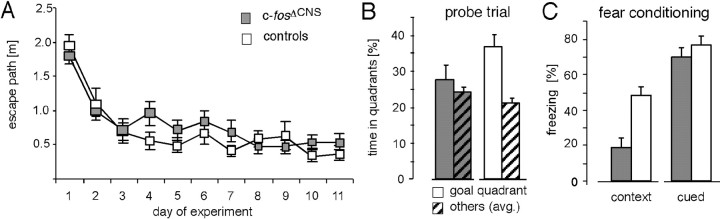
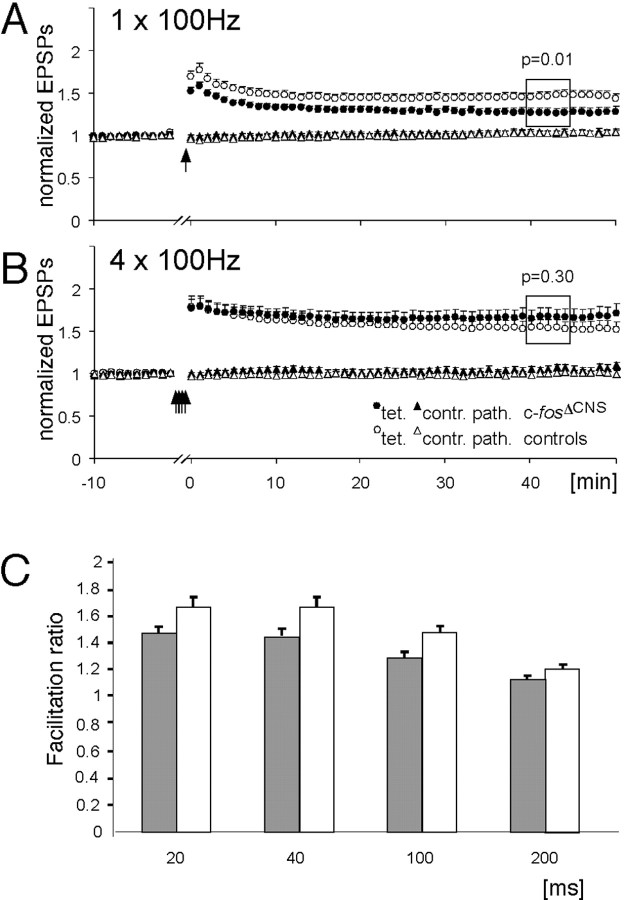
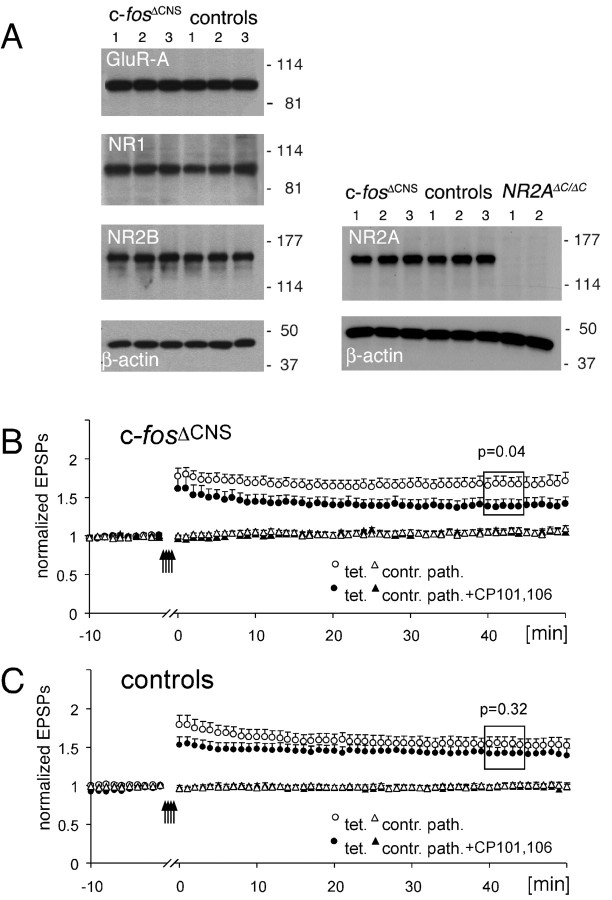
The immediate early gene c-fos is part of the activator protein-1 transcription factor and has been postulated to participate in the molecular mechanisms of learning and memory. To test this hypothesis in vivo, we generated mice with a nervous system-specific c-fos knock-out using the Cre-loxP system. Adult mice lacking c-Fos in the CNS (c-fosDeltaCNS) showed normal general and emotional behavior but were specifically impaired in hippocampus-dependent spatial and associative learning tasks. These learning deficits correlated with a reduction of long-term potentiation (LTP) in hippocampal CA3-CA1 synapses. The magnitude of LTP was restored by a repeated tetanization procedure, suggesting impaired LTP induction in c-fosDeltaCNS mice. This rescue was blocked by a selective inhibitor of NR2B-type NMDA receptors. This blockade was compensated in wild-type mice by NR2A-type NMDA receptor-activated signaling pathways, thus indicating that these pathways are compromised in c-fosDeltaCNS mice. In summary, our data suggest a role for c-Fos in hippocampus-dependent learning and memory as well as in NMDA receptor-dependent LTP formation.
Figures
References
-
- Andersen P ( 1960) Interhippocampal impulses. II. Apical dendritic activation of CA1 neurons. Acta Physiol Scand 48: 178-208. - PubMed
-
- Bannerman DM, Good MA, Butcher SP, Ramsay M, Morris RG ( 1995) Distinct components of spatial learning revealed by prior training and NMDA receptor blockade. Nature 378: 182-186. - PubMed
-
- Chiu R, Boyle WJ, Meek J, Smeal T, Hunter T, Karin M ( 1988) The c-Fos protein interacts with c-Jun/AP-1 to stimulate transcription of AP-1 responsive genes. Cell 54: 541-552. - PubMed
-
- Foster TC, McNaughton BL ( 1991) Long-term enhancement of CA1 synaptic transmission is due to increased quantal size, not quantal content. Hippocampus 1: 79-91. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous