

Loss of SEC-23 in Caenorhabditis elegans causes defects in oogenesis, morphogenesis, and extracellular matrix secretion
- PMID: 14551256
- PMCID: PMC266761
- DOI: 10.1091/mbc.e03-03-0162
Loss of SEC-23 in Caenorhabditis elegans causes defects in oogenesis, morphogenesis, and extracellular matrix secretion
Abstract
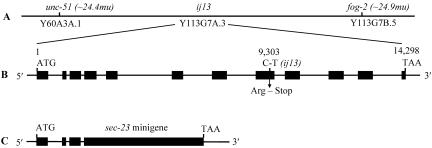
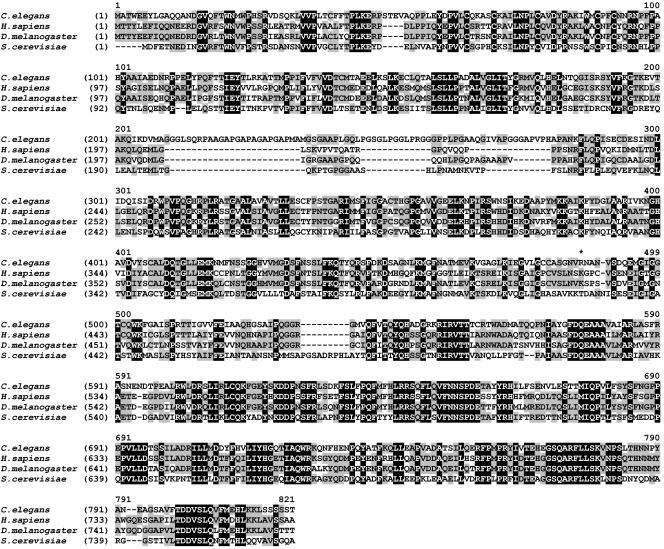
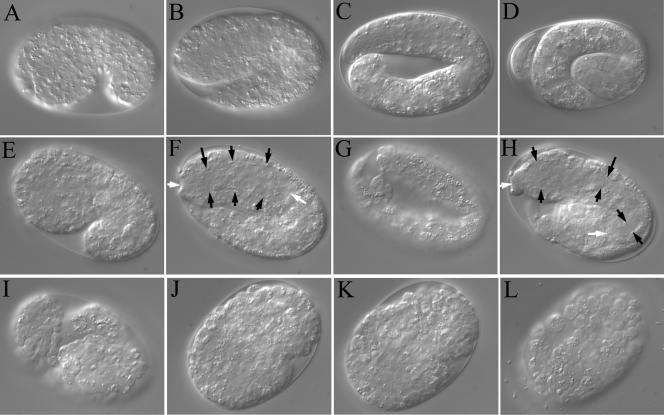
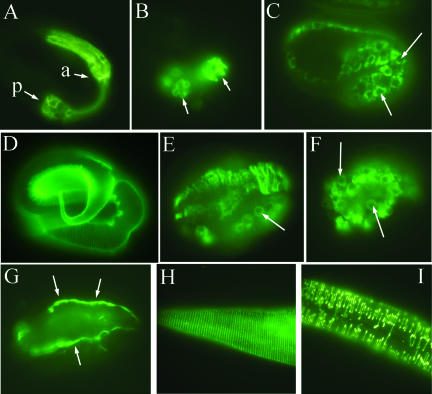
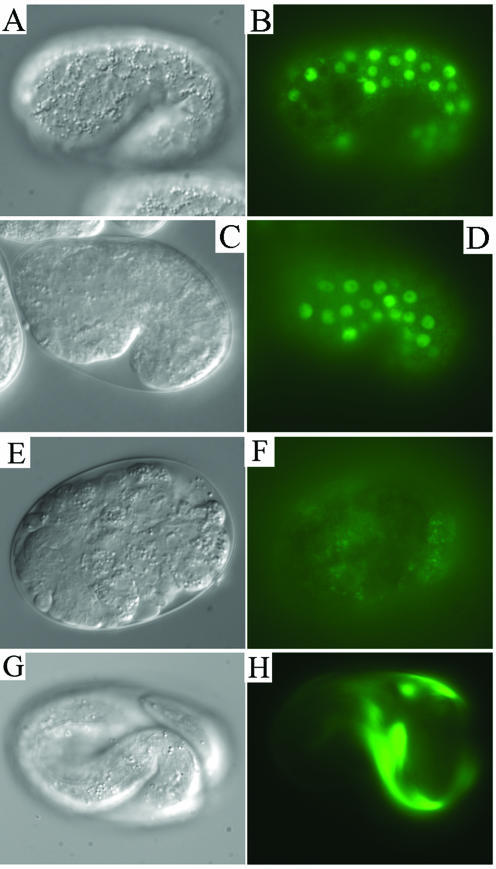
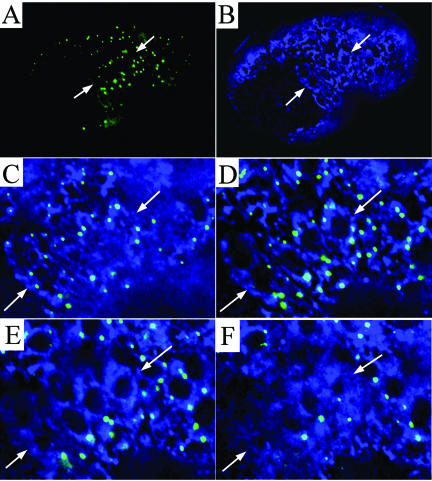
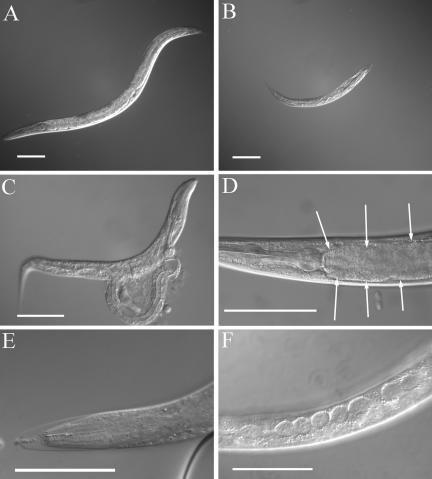
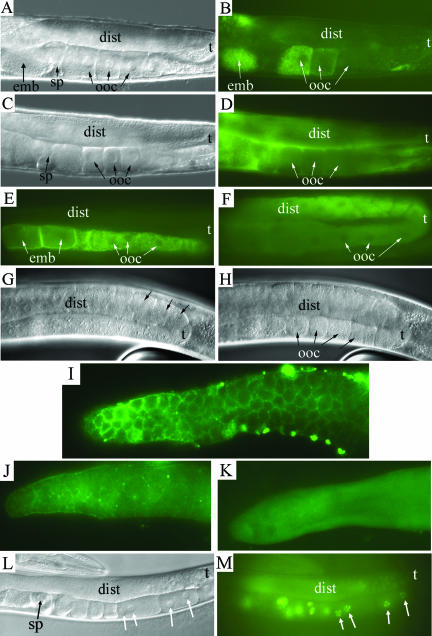
SEC-23 is a component of coat protein complex II (COPII)-coated vesicles involved in the endoplasmic reticulum-to-Golgi transport pathway of eukaryotes. During postembryonic life, Caenorhabditis elegans is surrounded by a collagenous exoskeleton termed the cuticle. From a screen for mutants defective in cuticle secretion, we identified and characterized a sec-23 mutant of C. elegans. By sequence homology, C. elegans has only the single sec-23 gene described herein. In addition to the cuticle secretion defect, mutants fail to complete embryonic morphogenesis. However, they progress through the earlier stages of embryogenesis, including gastrulation, and achieve substantial morphogenesis before death. We demonstrated a maternal component of SEC-23 function sufficient for progression through the earlier stages of embryogenesis and explaining the limited phenotype of the zygotic mutant. By RNA-mediated interference, we investigated the effects of perturbing COPII function during various postembryonic stages. During larval stages, major defects in cuticle synthesis and molting were observed. In the adult hermaphrodite, reduction of SEC-23 function by RNA-mediated interference caused a rapid onset of sterility, with defects in oogenesis including early maturation of the germline nuclei, probably a result of the observed loss of the GLP-1 receptor from the membrane surfaces adjacent to the developing germline nuclei.
Figures
Similar articles
-
The conserved transmembrane protein TMEM-39 coordinates with COPII to promote collagen secretion and regulate ER stress response.PLoS Genet. 2021 Feb 1;17(2):e1009317. doi: 10.1371/journal.pgen.1009317. eCollection 2021 Feb. PLoS Genet. 2021. PMID: 33524011 Free PMC article.
-
HRP-2, a heterogeneous nuclear ribonucleoprotein, is essential for embryogenesis and oogenesis in Caenorhabditis elegans.Exp Cell Res. 2004 Aug 15;298(2):418-30. doi: 10.1016/j.yexcr.2004.04.043. Exp Cell Res. 2004. PMID: 15265690
-
Combined extracellular matrix cross-linking activity of the peroxidase MLT-7 and the dual oxidase BLI-3 is critical for post-embryonic viability in Caenorhabditis elegans.J Biol Chem. 2009 Jun 26;284(26):17549-63. doi: 10.1074/jbc.M900831200. Epub 2009 Apr 30. J Biol Chem. 2009. PMID: 19406744 Free PMC article.
-
Vesicle-mediated export from the ER: COPII coat function and regulation.Biochim Biophys Acta. 2013 Nov;1833(11):2464-72. doi: 10.1016/j.bbamcr.2013.02.003. Epub 2013 Feb 15. Biochim Biophys Acta. 2013. PMID: 23419775 Free PMC article. Review.
-
COPII and the regulation of protein sorting in mammals.Nat Cell Biol. 2011 Dec 22;14(1):20-8. doi: 10.1038/ncb2390. Nat Cell Biol. 2011. PMID: 22193160 Review.
Cited by
-
Extracellular leucine-rich repeat proteins are required to organize the apical extracellular matrix and maintain epithelial junction integrity in C. elegans.Development. 2012 Mar;139(5):979-90. doi: 10.1242/dev.075135. Epub 2012 Jan 25. Development. 2012. PMID: 22278925 Free PMC article.
-
Functional genomic analysis of C. elegans molting.PLoS Biol. 2005 Oct;3(10):e312. doi: 10.1371/journal.pbio.0030312. Epub 2005 Aug 30. PLoS Biol. 2005. PMID: 16122351 Free PMC article.
-
Genetics coupled to quantitative intact proteomics links heritable aphid and endosymbiont protein expression to circulative polerovirus transmission.J Virol. 2011 Mar;85(5):2148-66. doi: 10.1128/JVI.01504-10. Epub 2010 Dec 15. J Virol. 2011. PMID: 21159868 Free PMC article.
-
The genetic basis of a craniofacial disease provides insight into COPII coat assembly.Dev Cell. 2007 Nov;13(5):623-634. doi: 10.1016/j.devcel.2007.10.005. Dev Cell. 2007. PMID: 17981132 Free PMC article.
-
Mechanistic insights and identification of two novel factors in the C. elegans NMD pathway.Genes Dev. 2007 May 1;21(9):1075-85. doi: 10.1101/gad.417707. Epub 2007 Apr 16. Genes Dev. 2007. PMID: 17437990 Free PMC article.
References
-
- Albertson, D.G., and Thomson, J.N. (1976). The pharynx of Caenorhabditis elegans. Phil. Trans. R. Soc. Lond. B Biol. Sci. 275, 299-325. - PubMed
-
- Anderson, P. (1995). Mutagenesis. In: Caenorhabditis elegans: Modern Biological Analysis of an Organism, ed. H.F. Epstein and D.C. Shakes, San Diego: Academic Press, 31-58.
-
- Barlowe, C., Orci, L., Yeung, T., Hosobuchi, M., Hamamoto, S., Salama, N., Rexach, M.F., Ravazzola, M., Amherdt, M., and Schekman, R. (1994). COPII a membrane coat formed by SEC proteins that drive vesicle budding from the endoplasmic reticulum. Cell 77, 895-907. - PubMed
-
- Bednarek, S.Y., Orci, L., and Schekman, R. (1996). Traffic COPs and the formation of vesicle coats. Trends Cell Biol. 6, 468-473. - PubMed
-
- Berry, L.W., Westlund, B., and Schedl, T. (1997). Germ-line tumor formation caused by activation of glp-1, a Caenorhabditis elegans member of the Notch family of receptors. Development 124, 925-936. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
