

Microspore separation in the quartet 3 mutants of Arabidopsis is impaired by a defect in a developmentally regulated polygalacturonase required for pollen mother cell wall degradation
- PMID: 14551328
- PMCID: PMC281612
- DOI: 10.1104/pp.103.028266
Microspore separation in the quartet 3 mutants of Arabidopsis is impaired by a defect in a developmentally regulated polygalacturonase required for pollen mother cell wall degradation
Abstract
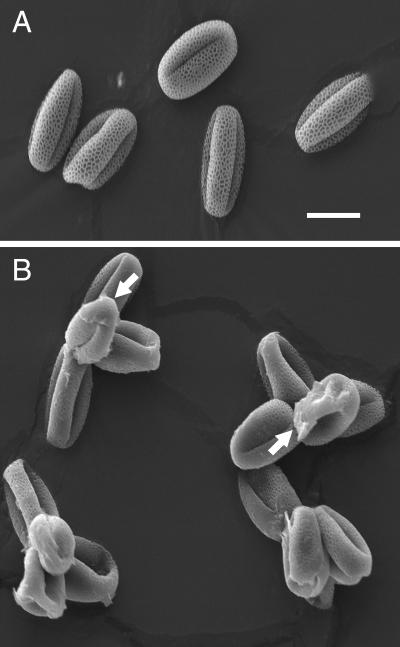
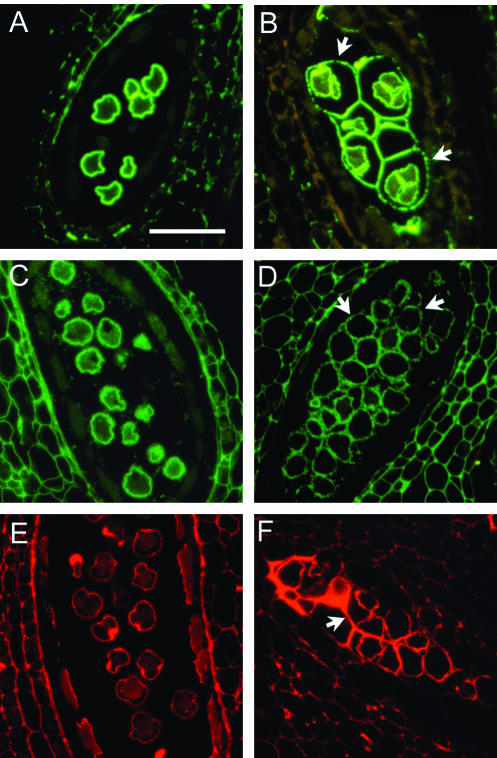
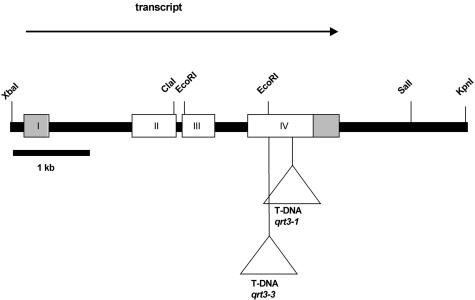
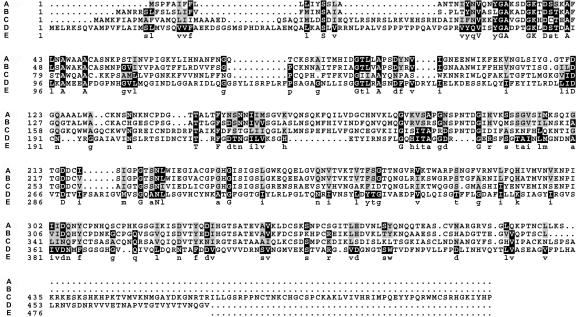
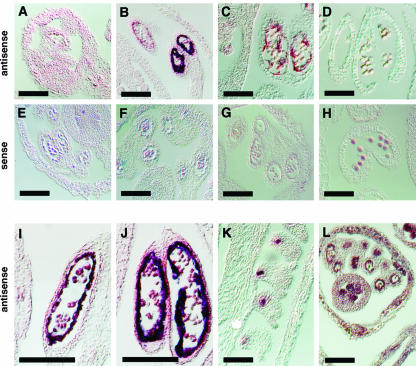
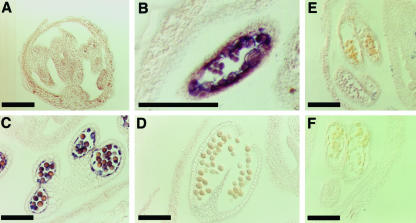
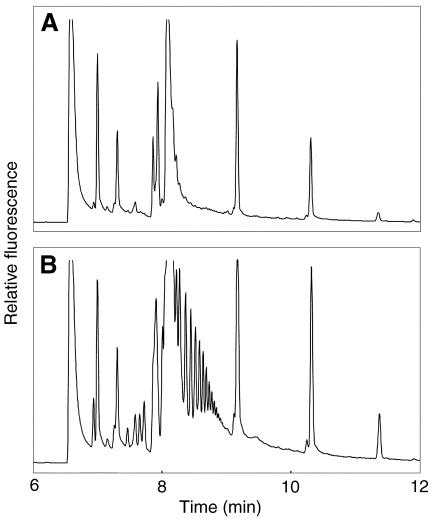
Mutations in the QUARTET loci in Arabidopsis result in failure of microspore separation during pollen development due to a defect in degradation of the pollen mother cell wall during late stages of pollen development. Mutations in a new locus required for microspore separation, QRT3, were isolated, and the corresponding gene was cloned by T-DNA tagging. QRT3 encodes a protein that is approximately 30% similar to an endopolygalacturonase from peach (Prunus persica). The QRT3 protein was expressed in yeast (Saccharomyces cerevisiae) and found to exhibit polygalacturonase activity. In situ hybridization experiments showed that QRT3 is specifically and transiently expressed in the tapetum during the phase when microspores separate from their meiotic siblings. Immunohistochemical localization of QRT3 indicated that the protein is secreted from tapetal cells during the early microspore stage. Thus, QRT3 plays a direct role in degrading the pollen mother cell wall during microspore development.
Figures

References
-
- Albani D, Altosaar I, Arnison PG, Fabijanski SF (1991) A gene showing sequence similarity to pectin esterase is specifically expressed in developing pollen of Brassica napus: sequences in its 5′ flanking region are conserved in other pollen-specific promoters. Plant Mol Biol 16: 501–513 - PubMed
-
- Amagai M, Ariizumi T, Endo M, Hatakeyama K, Kuwata C, Shibata D, Toriyama K, Watanabe M (2003) Identification of anther-specific genes in a cruciferous model plant, Arabidopsis thaliana, by using a combination of Arabidopsis macroarray and mRNA derived from Brassica oleracea. Sex Plant Reprod 15: 213–220
-
- Ariizumi T, Amagai M, Shibata D, Hatakeyama K, Watanabe M, Toriyama K (2002) Comparative study of promoter activity of three anther-specific genes encoding lipid transfer protein, xyloglucan endotransglucosylase/hydrolase and polygalacturonase in transgenic Arabidopsis thaliana. Plant Cell Rep 21: 90–96
-
- Ausubel FM, Brent R, Kingston RE, Moore DD, Seidman JG, Smith JA, Struhl K (1989) Preparation of genomic DNA from plant tissue. In FM Ausubel, R Bent, RE Kingston, DD Moore, JG Seidman, JA Smith, K Struhl, eds, Current Protocols in Molecular Biology, Vol. 1. Wiley, New York, pp 2.3.1–2.3.3
-
- Barnett MJ, Long SR (1997) Identification and characterization of a gene on Rhizobium meliloti pSyma, syrB, that negatively affects syrM expression. Mol Plant-Microbe Interact 10: 550–559 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
