

Exposure to low infective doses of HCV induces cellular immune responses without consistently detectable viremia or seroconversion in chimpanzees
- PMID: 14554088
- PMCID: PMC3898342
- DOI: 10.1016/s0042-6822(03)00461-6
Exposure to low infective doses of HCV induces cellular immune responses without consistently detectable viremia or seroconversion in chimpanzees
Abstract
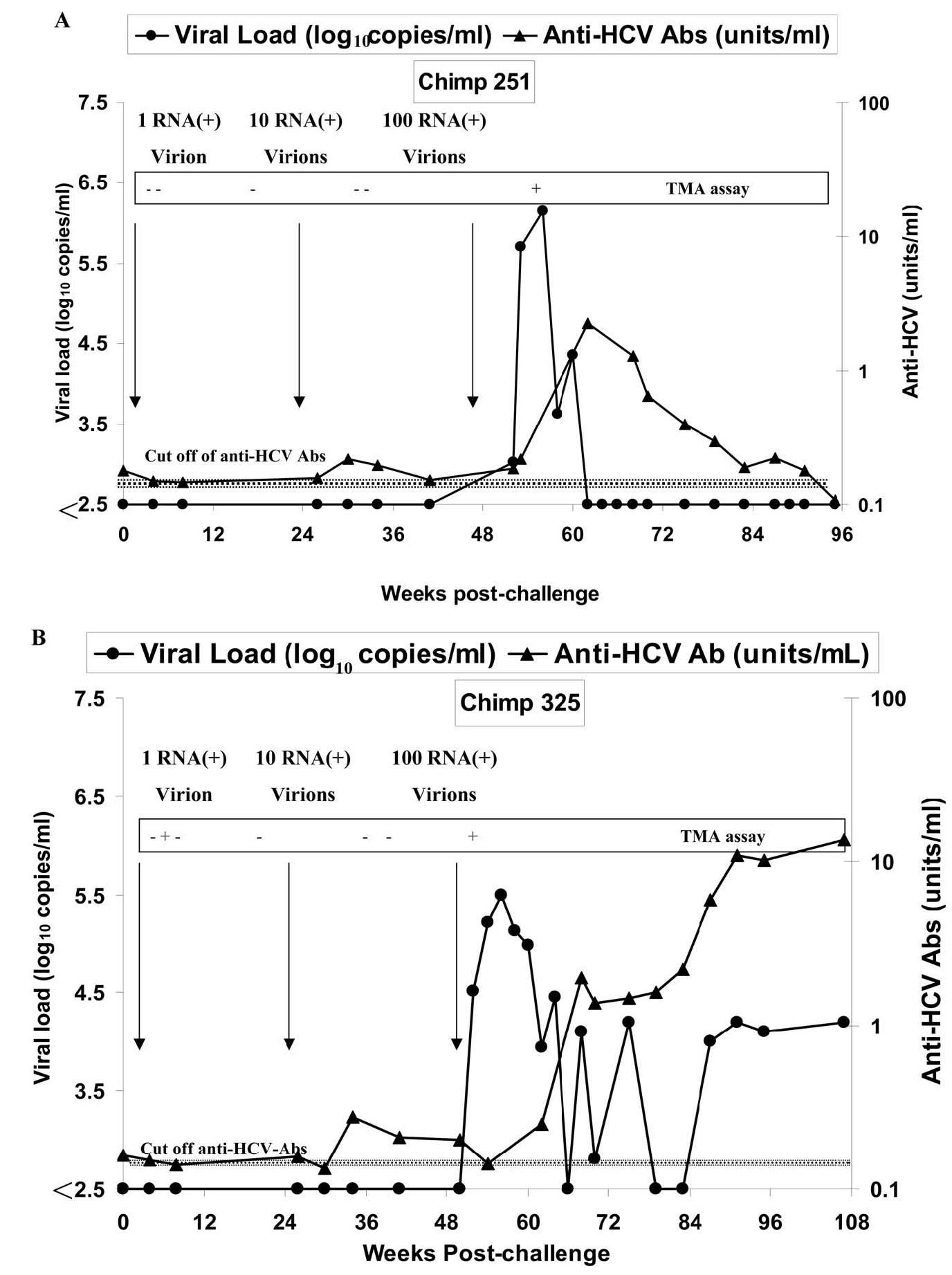
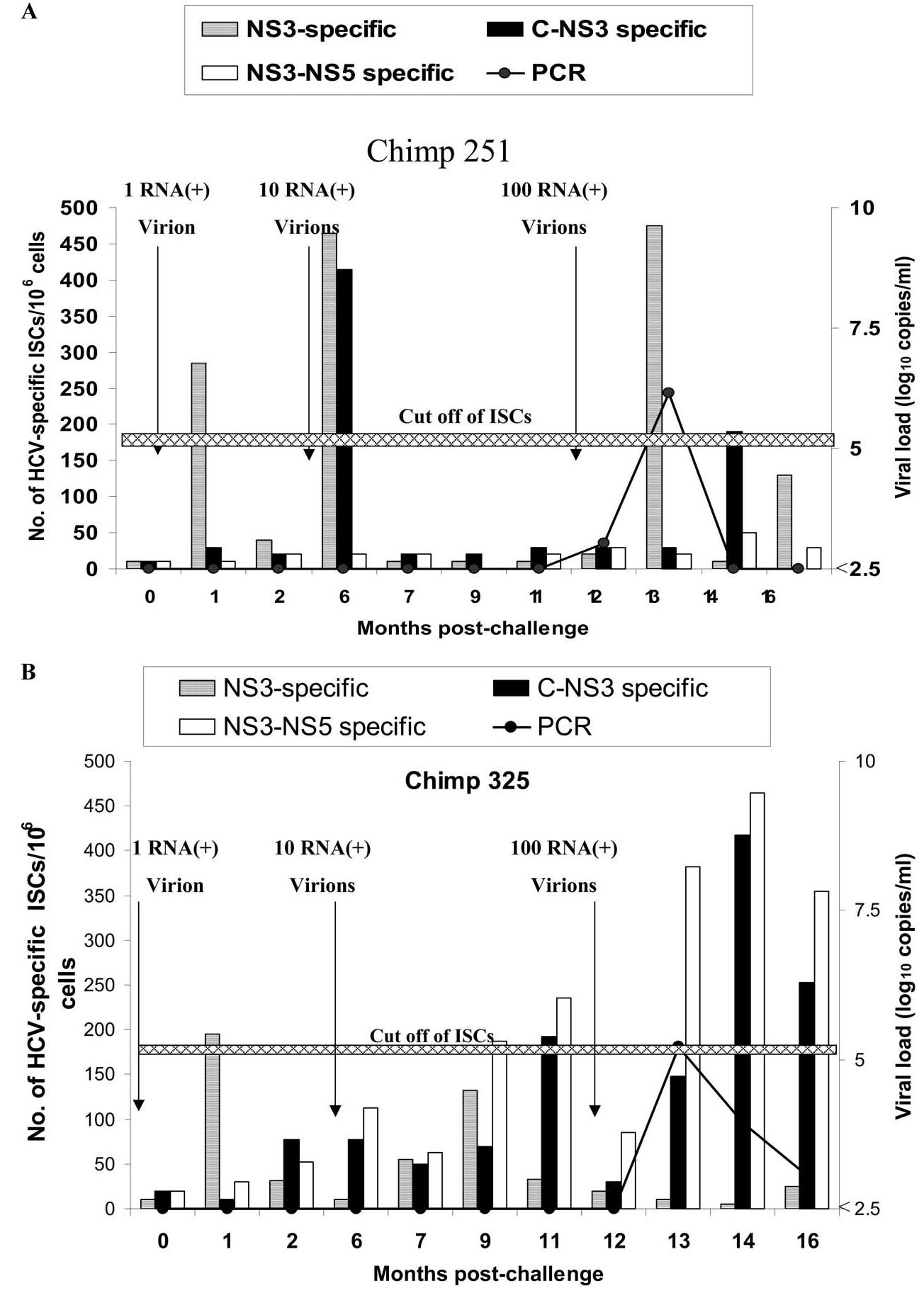
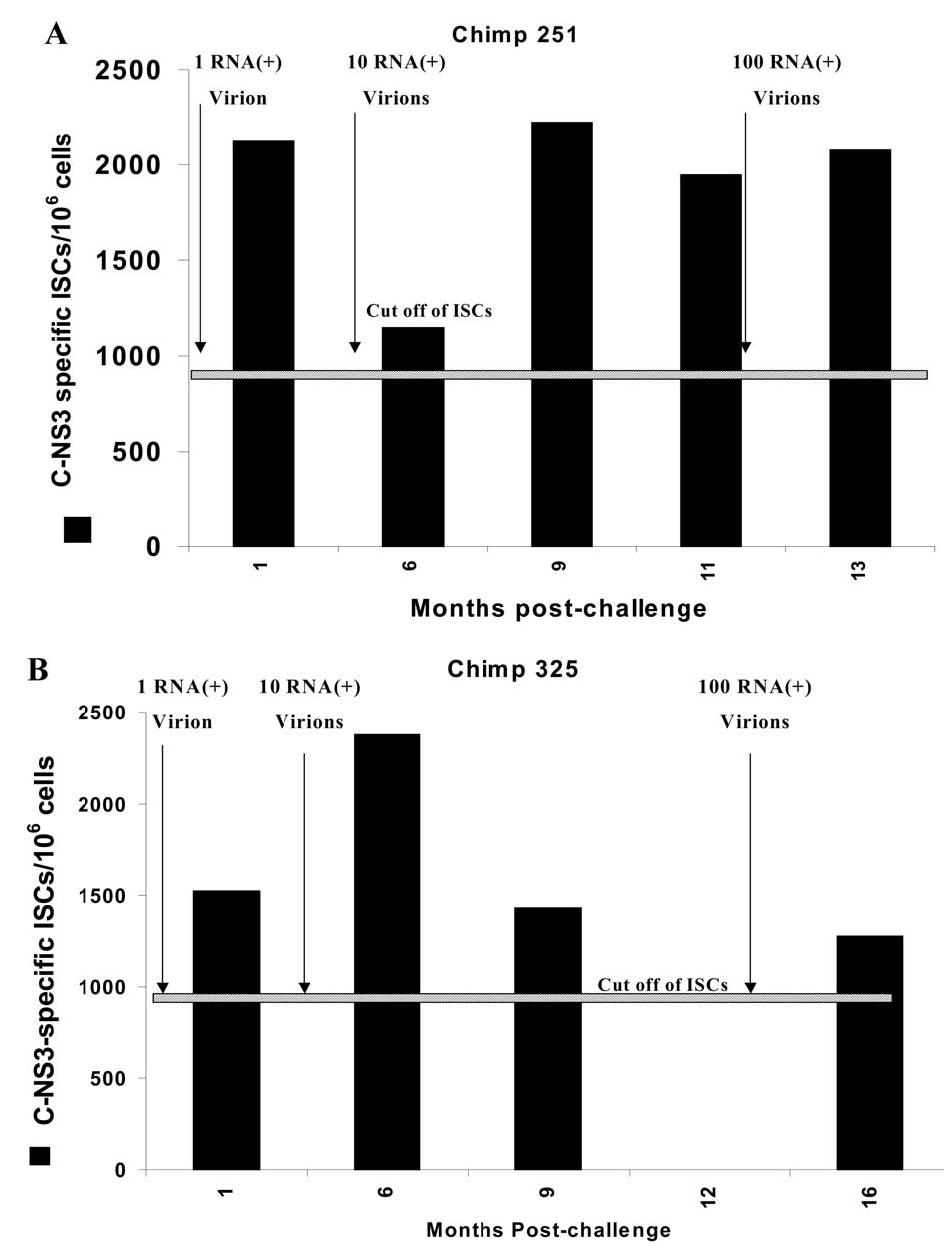
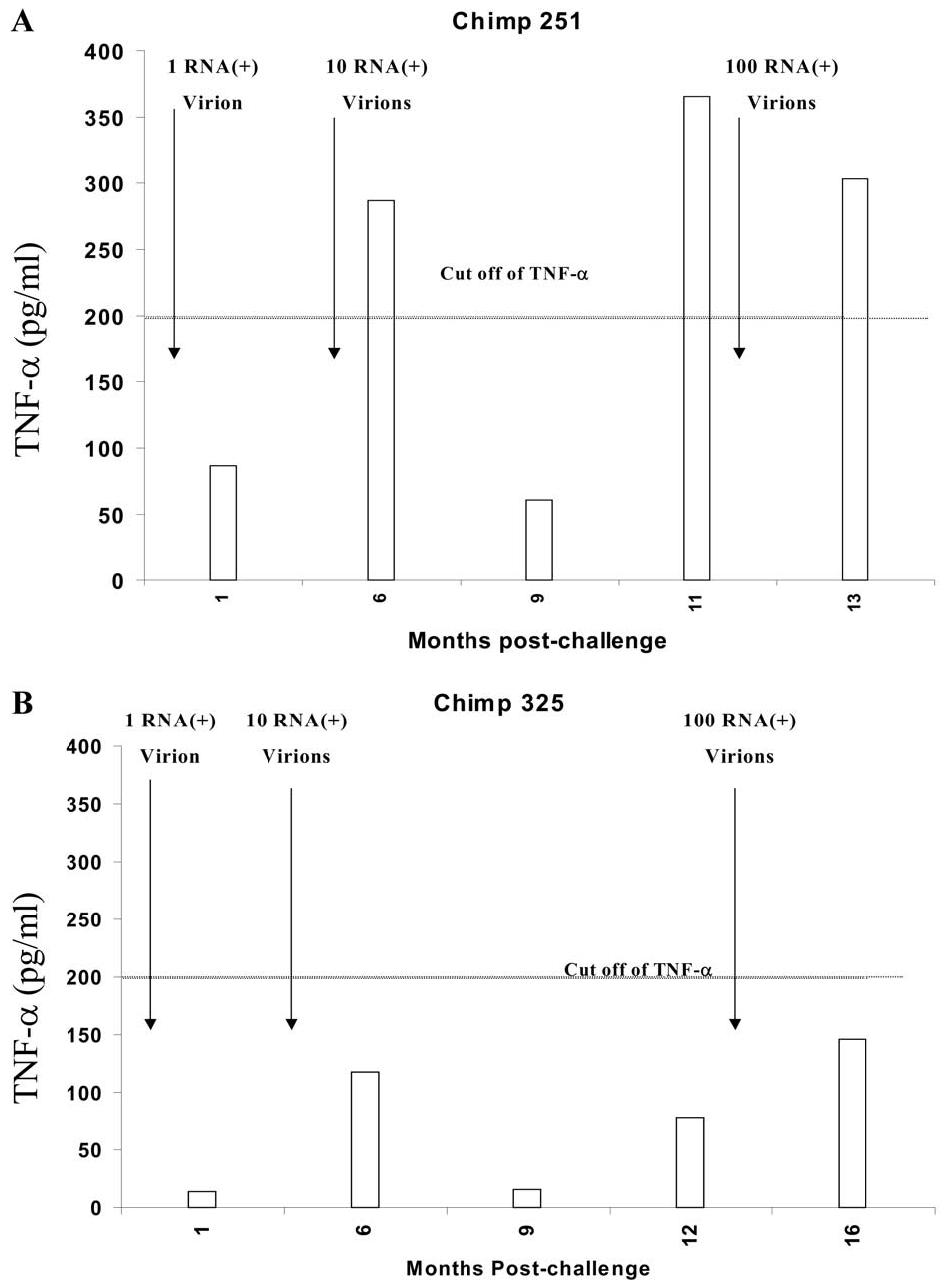
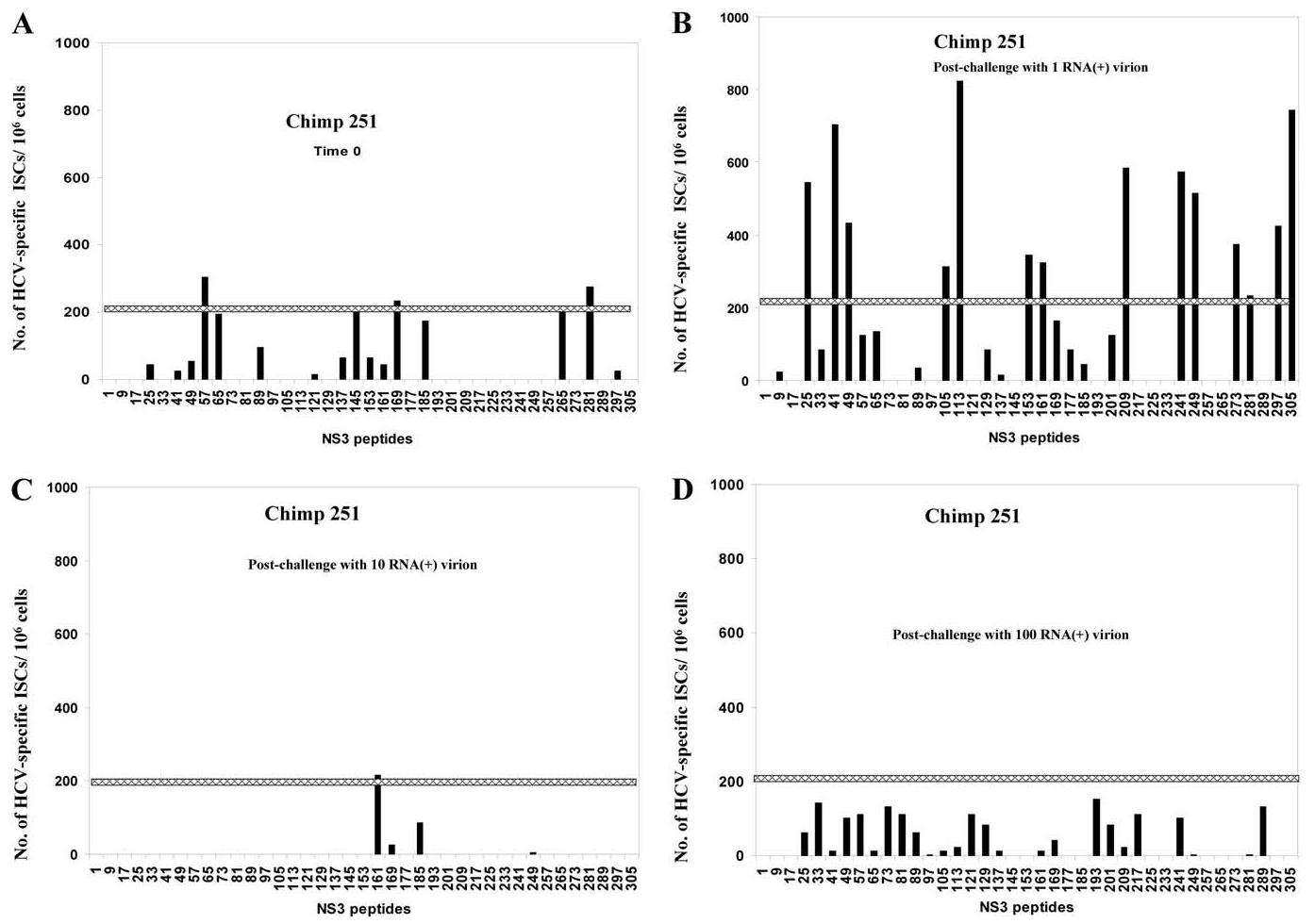
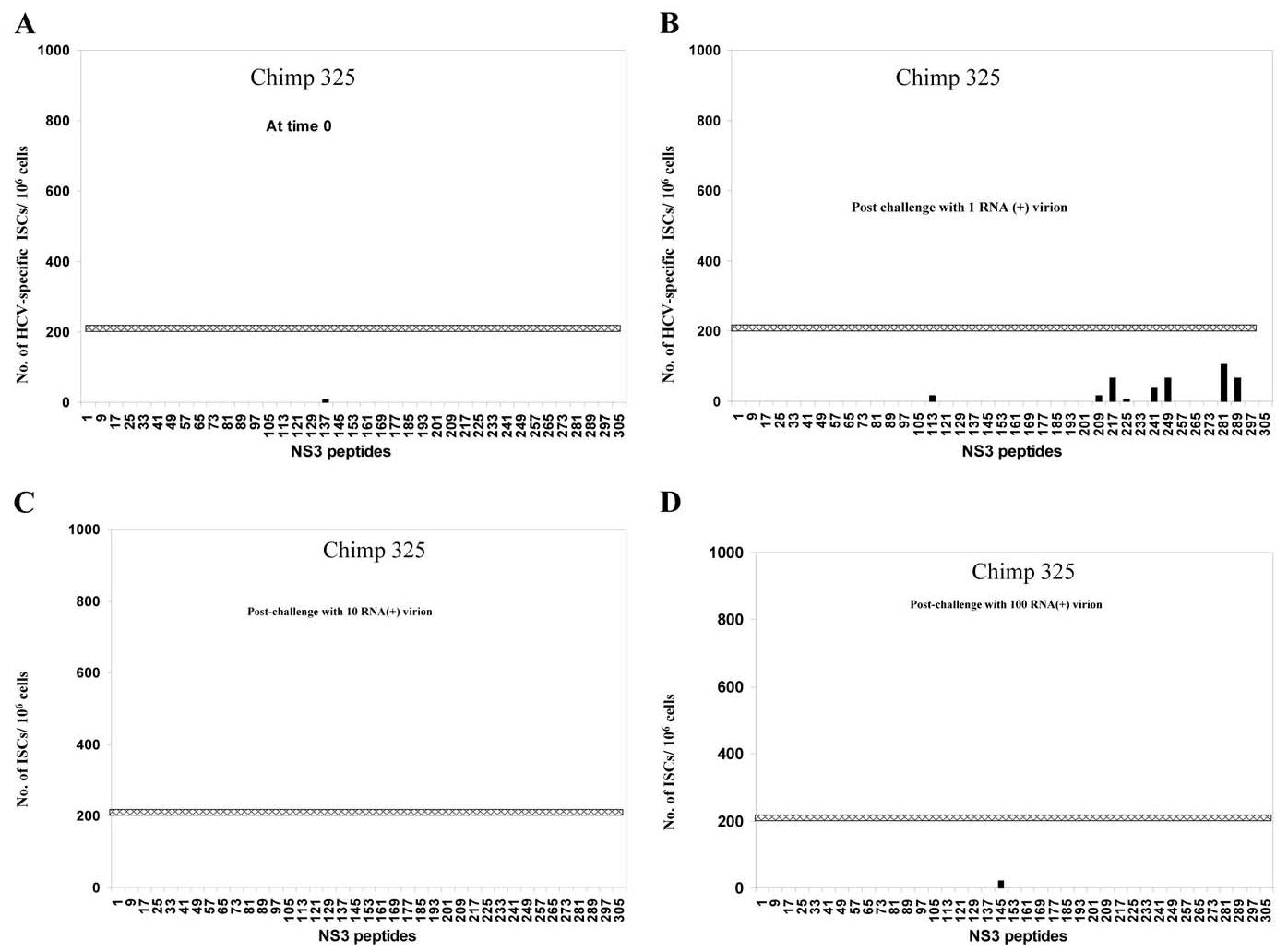
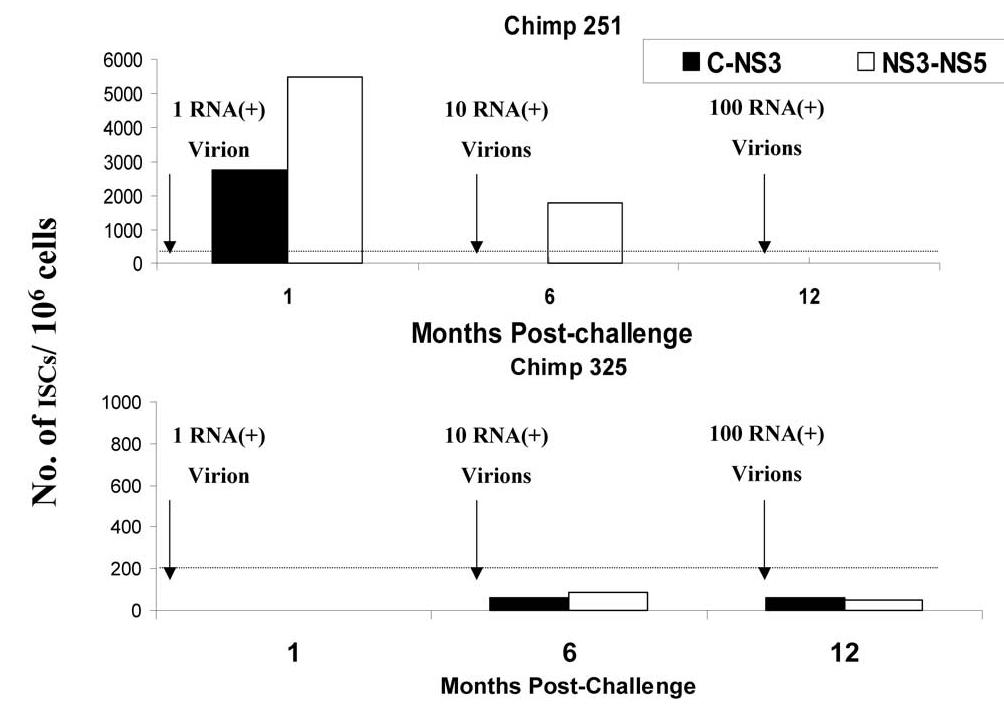
In hepatitis C virus (HCV) infection, there is accumulating data suggesting the presence of cellular immune responses to HCV in exposed but seemingly uninfected populations. Some studies have suggested cross-reactive antigens rather than prior HCV exposure as the main reason for the immune responses. In this study we address this question by analyzing the immune response of chimpanzees that have been sequentially exposed to increasing doses of HCV virions. The level of viremia, as well as the immune responses to HCV at different times after virus inoculation, were examined. Our data indicate that HCV infective doses as low as 1-10 RNA (+) virions induce detectable cellular immune responses in chimpanzees without consistently detectable viremia or persistent seroconversion. However, increasing the infective doses of HCV to 100 RNA (+) virions overcame the low-inoculum-induced immune response and produced high-level viremia followed by seroconversion.
Figures
Similar articles
-
Kinetics of CD4+ and CD8+ memory T-cell responses during hepatitis C virus rechallenge of previously recovered chimpanzees.J Virol. 2003 Apr;77(8):4781-93. doi: 10.1128/jvi.77.8.4781-4793.2003. J Virol. 2003. PMID: 12663785 Free PMC article.
-
Previously infected and recovered chimpanzees exhibit rapid responses that control hepatitis C virus replication upon rechallenge.J Virol. 2002 Jul;76(13):6586-95. doi: 10.1128/jvi.76.13.6586-6595.2002. J Virol. 2002. PMID: 12050371 Free PMC article.
-
Clearance of low levels of HCV viremia in the absence of a strong adaptive immune response.Virol J. 2007 Jun 11;4:58. doi: 10.1186/1743-422X-4-58. Virol J. 2007. PMID: 17562015 Free PMC article.
-
The role of immune responses in the pathogenesis of hepatitis C virus infection.J Viral Hepat. 1997;4 Suppl 2:31-41. doi: 10.1111/j.1365-2893.1997.tb00178.x. J Viral Hepat. 1997. PMID: 9429208 Review.
-
Cell-mediated immunity and the outcome of hepatitis C virus infection.Annu Rev Microbiol. 2004;58:391-424. doi: 10.1146/annurev.micro.58.030603.123836. Annu Rev Microbiol. 2004. PMID: 15487943 Review.
Cited by
-
Cellular immune responses in seronegative sexual contacts of acute hepatitis C patients.J Virol. 2004 Nov;78(22):12252-8. doi: 10.1128/JVI.78.22.12252-12258.2004. J Virol. 2004. PMID: 15507612 Free PMC article.
-
Innate immune responses in hepatitis C virus-exposed healthcare workers who do not develop acute infection.Hepatology. 2013 Nov;58(5):1621-31. doi: 10.1002/hep.26353. Epub 2013 Sep 30. Hepatology. 2013. PMID: 23463364 Free PMC article.
-
Hepatitis C virus-multispecific T-cell responses without viremia or seroconversion among Egyptian health care workers at high risk of infection.Clin Vaccine Immunol. 2012 May;19(5):780-6. doi: 10.1128/CVI.00050-12. Epub 2012 Mar 21. Clin Vaccine Immunol. 2012. PMID: 22441392 Free PMC article.
-
Risk factors and immune response to hepatitis E viral infection among acute hepatitis patients in Assiut, Egypt.Egypt J Immunol. 2010;17(1):73-86. Egypt J Immunol. 2010. PMID: 22053611 Free PMC article.
-
Two distinct functional patterns of hepatitis C Virus (HCV)-specific T cell responses in seronegative, aviremic patients.PLoS One. 2013 Apr 30;8(4):e62319. doi: 10.1371/journal.pone.0062319. Print 2013. PLoS One. 2013. PMID: 23638039 Free PMC article.
References
-
- Abe K, Inchauspe G, Shikata T, Prince AM. Three different patterns of hepatitis C virus infection in chimpanzees. Hepatology. 1992;15((4)):690–695. - PubMed
-
- Alter MJ, Kruszon-Moran D, Nainan OV, McQuillan GM, Gao F, Moyer LA, Kaslow RA, Margolis HS. The prevalence of hepatitis C virus infection in the United States, 1988 through 1994. N. Engl. J. Med. 1999;341((8)):556–562. - PubMed
-
- Ando K, Hiroishi K, Kaneko T, Moriyama T, Muto Y, Kayagaki N, Yagita H, Okumura K, Imawari M. Perforin, Fas/Fas ligand, and TNF-alpha pathways as specific and bystander killing mechanisms of hepatitis C virus-specific human CTL. J. Immunol. 1997;158((11)):5283–5291. - PubMed
-
- Anthony DD, Post AB, Valdez H, Peterson DL, Murphy M, Heeger PS. ELISPOT analysis of hepatitis C virus protein-specific IFN-gamma-producing peripheral blood lymphocytes in infected humans with and without cirrhosis. Clin. Immunol. 2001;99((2)):232–240. - PubMed
-
- Bassett SE, Guerra B, Brasky K, Miskovsky E, Houghton M, Klimpel GR, Lanford RE. Protective immune response to hepatitis C virus in chimpanzees rechallenged following clearance of primary infection. Hepatology. 2001;33((6)):1479–1487. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
