

The modulation by 5-HT of glutamatergic inputs from the raphe pallidus to rat hypoglossal motoneurones, in vitro
- PMID: 14555716
- PMCID: PMC2343612
- DOI: 10.1113/jphysiol.2003.053843
The modulation by 5-HT of glutamatergic inputs from the raphe pallidus to rat hypoglossal motoneurones, in vitro
Abstract
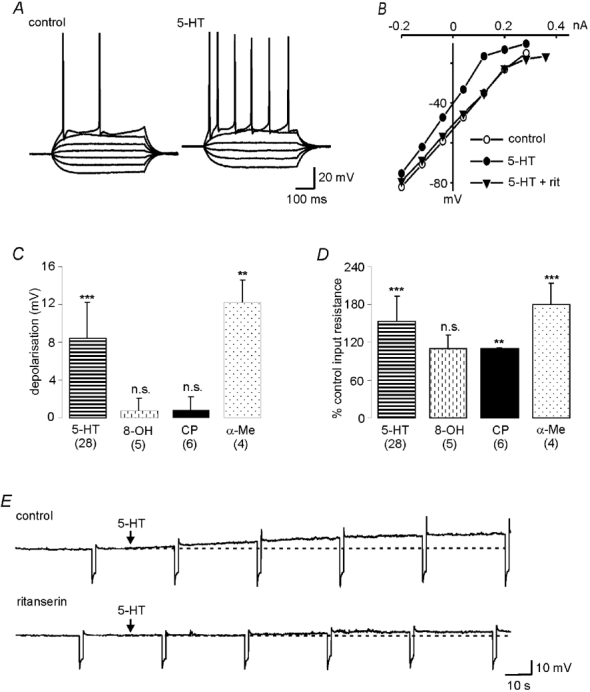
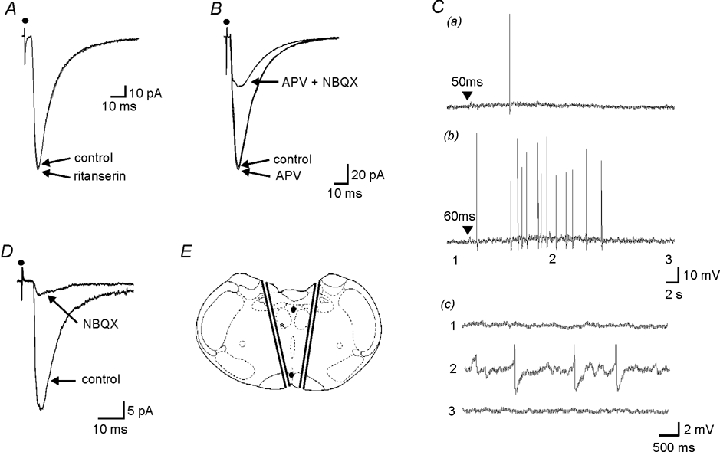
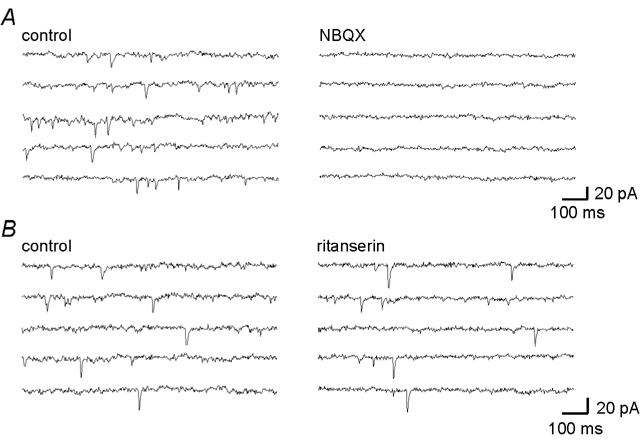
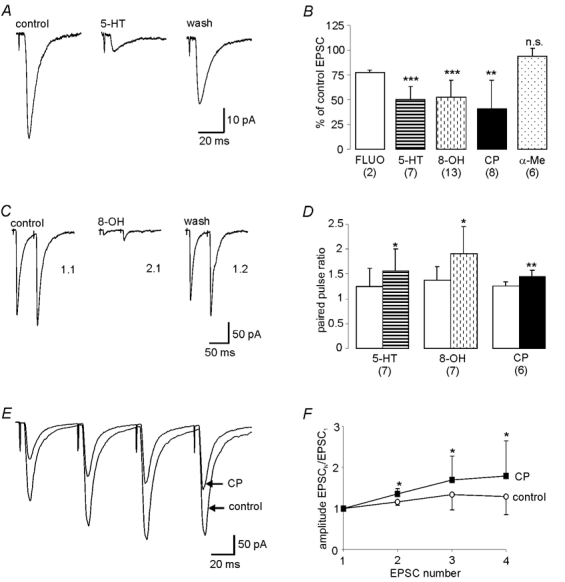
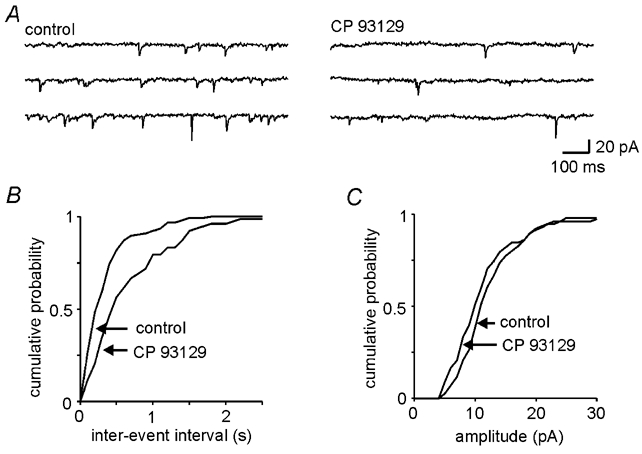
Decreases in the activity of 5-HT-containing caudal raphe neurones during sleep are thought to be partially responsible for the resultant disfacilitation of hypoglossal motoneurones. Whilst 5-HT has a direct excitatory action on hypoglossal motoneurones as a result of activation of 5-HT2 receptors, microinjection of 5-HT2 antagonists into the hypoglossal nucleus reduces motor activity to a much lesser extent compared to the suppression observed during sleep suggesting other transmitters co-localised in caudal raphe neurones may also be involved. The aim of the present study was therefore to characterise raphe pallidus inputs to hypoglossal motoneurones. Whole cell recordings were made from hypoglossal motoneurones in vitro. 5-HT evoked a direct membrane depolarisation (8.45 +/- 3.8 mV, P < 0.001) and increase in cell input resistance (53 +/- 40 %, P < 0.001) which was blocked by the 5-HT2 antagonist, ritanserin (2.40 +/- 2.7 vs. 7.04 +/- 4.6 mV). Stimulation within the raphe pallidus evoked a monosynaptic EPSC that was significantly reduced by the AMPA/kainate antagonist, NBQX (22.8 +/- 16 % of control, P < 0.001). In contrast, the 5-HT2 antagonist, ritanserin, had no effect on the amplitude of these EPSCs (106 +/- 31 % of control, P = n.s.). 5-HT reduced these EPSCs to 50.0 +/- 13 % of control (P < 0.001), as did the 5-HT1A agonist, 8-OH-DPAT (52.5 +/- 17 %, P < 0.001) and the 5-HT1B agonist, CP 93129 (40.6 +/- 29 %, P < 0.01). 8-OH-DPAT and CP 93129 increased the paired pulse ratio (1.38 +/- 0.27 to 1.91 +/- 0.54, P < 0.05 & 1.27 +/- 0.08 to 1.44 +/- 0.13, P < 0.01 respectively) but had no effect on the postsynaptic glutamate response (99 +/- 4.4 % and 100 +/- 2.5 %, P = n.s.). They also increased the frequency (P < 0.001), but not the amplitude, of miniature glutamatergic EPSCs in hypoglossal motoneurones. These data demonstrate that raphe pallidus inputs to hypoglossal motoneurones are predominantly glutamatergic in nature, with 5-HT decreasing the release of glutamate from these projections as a result of activation of 5-HT1A and/or 5-HT1B receptors located on presynaptic terminals.
Figures
References
-
- Aldes LD, Marco LA, Chronister RB. Serotonin-containing axon terminals in the hypoglossal nucleus of the rat. An immuno-electronmicroscopic study. Brain Res Bull. 1989;23:249–256. - PubMed
-
- Al-Zubaidy ZA, Erickson RL, Greer JJ. Serotonergic and noradrenergic effects on respiratory neural discharge in the medullary slice preparation of neonatal rats. Eur J Physiol. 1996;431:942–949. - PubMed
-
- Arita H, Ochiishi M. Opposing effects of 5-HT on two types of medullary inspiratory neurones with distinct firing patterns. J Neurophysiol. 1991;66:285–292. - PubMed
-
- Bayliss DA, Viana F, Talley EM, Berger AJ. Neuromodulation of hypoglossal motoneurons: cellular and developmental mechanisms. Respir Physiol. 1997;110:139–150. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous