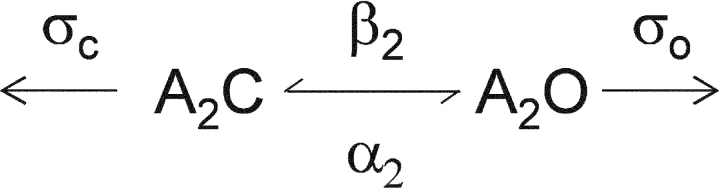The role of loop 5 in acetylcholine receptor channel gating
- PMID: 14557402
- PMCID: PMC2229574
- DOI: 10.1085/jgp.200308885
The role of loop 5 in acetylcholine receptor channel gating
Abstract
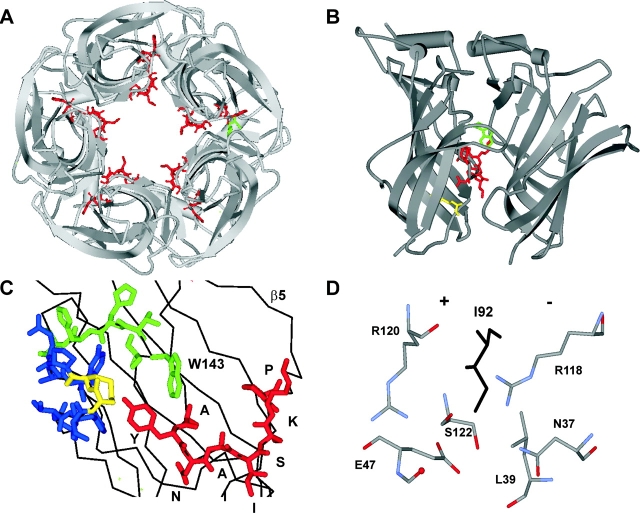
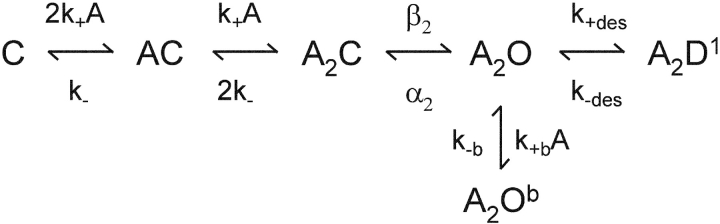
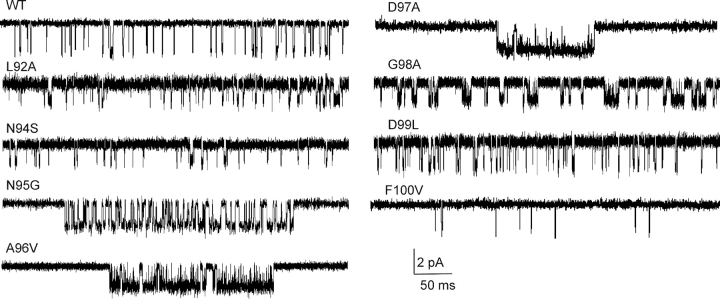
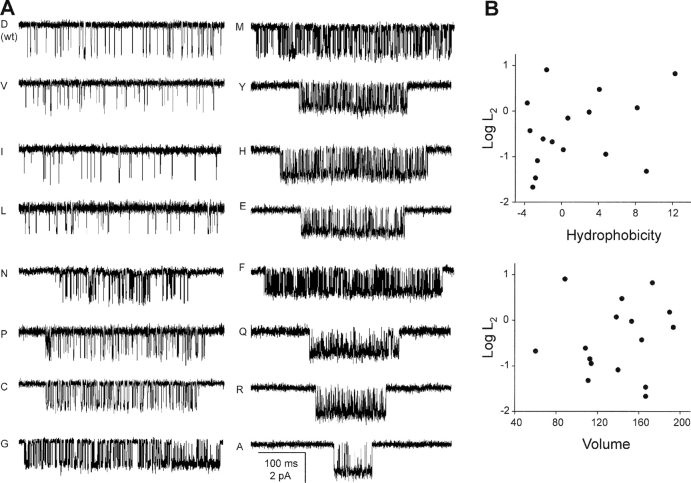
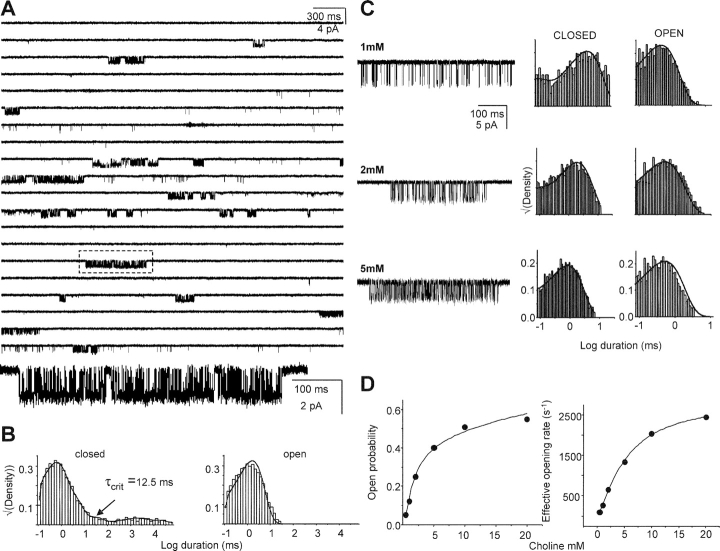
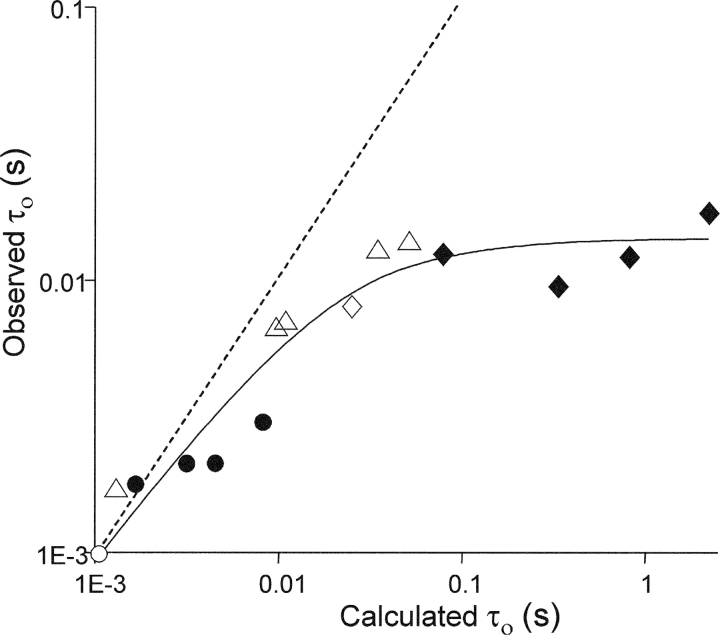
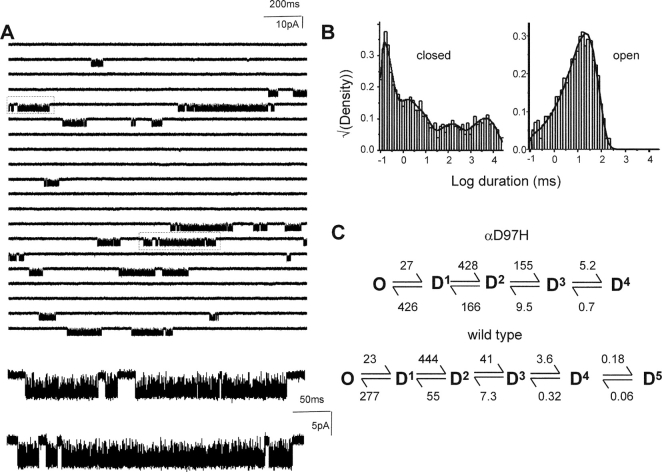
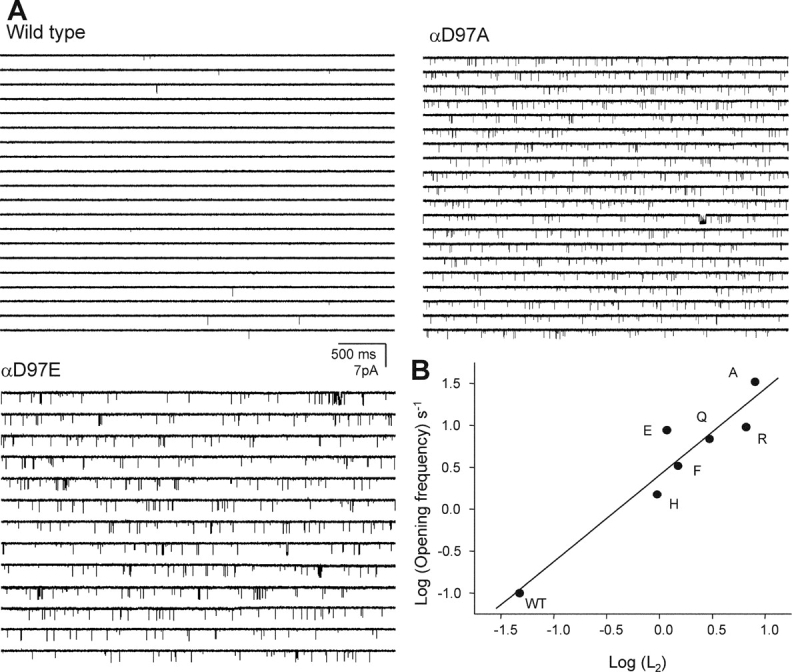
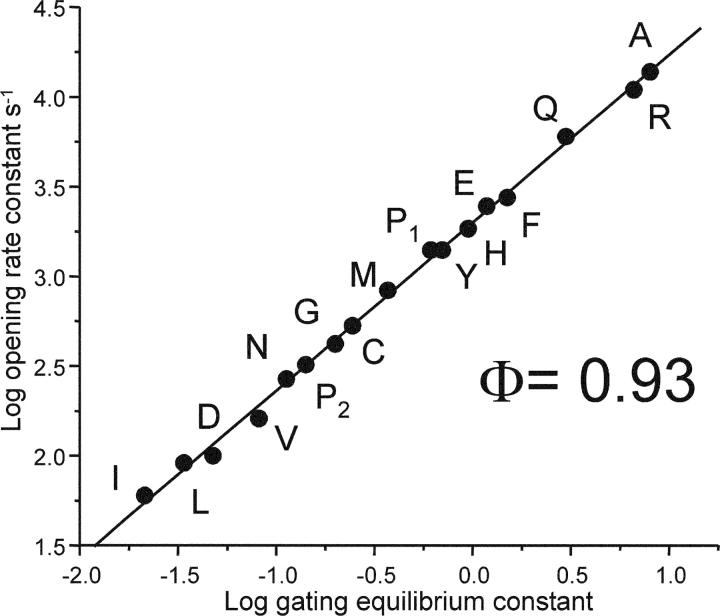
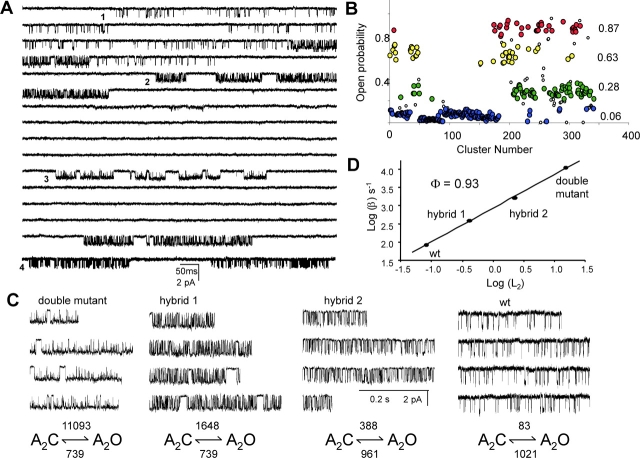
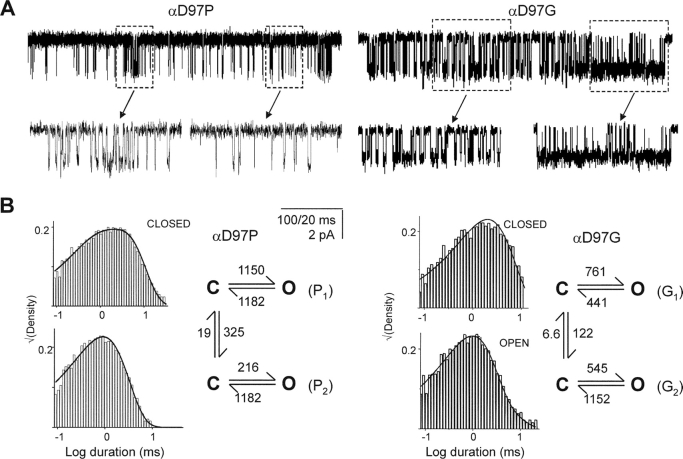
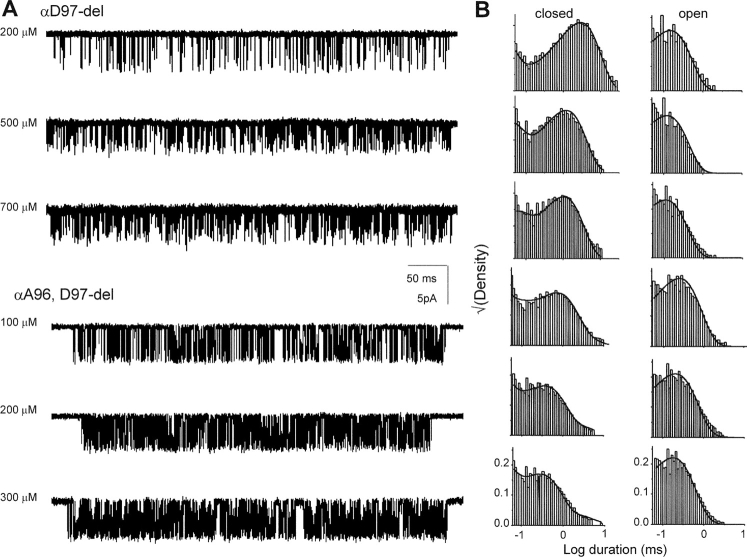
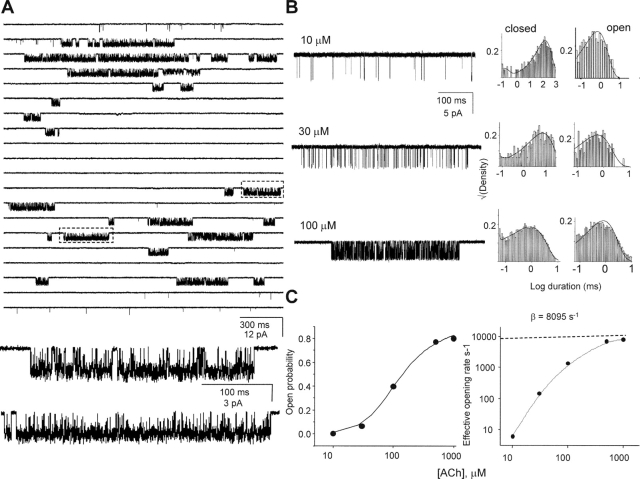
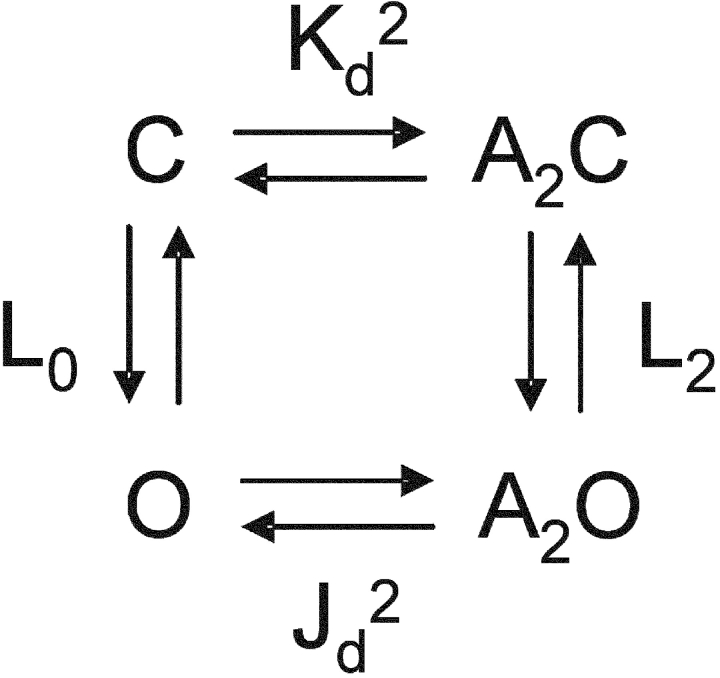
Nicotinic acetylcholine receptor channel (AChR) gating is an organized sequence of molecular motions that couples a change in the affinity for ligands at the two transmitter binding sites with a change in the ionic conductance of the pore. Loop 5 (L5) is a nine-residue segment (mouse alpha-subunit 92-100) that links the beta4 and beta5 strands of the extracellular domain and that (in the alpha-subunit) contains binding segment A. Based on the structure of the acetylcholine binding protein, we speculate that in AChRs L5 projects from the transmitter binding site toward the membrane along a subunit interface. We used single-channel kinetics to quantify the effects of mutations to alphaD97 and other L5 residues with respect to agonist binding (to both open and closed AChRs), channel gating (for both unliganded and fully-liganded AChRs), and desensitization. Most alphaD97 mutations increase gating (up to 168-fold) but have little or no effect on ligand binding or desensitization. Rate-equilibrium free energy relationship analysis indicates that alphaD97 moves early in the gating reaction, in synchrony with the movement of the transmitter binding site (Phi = 0.93, which implies an open-like character at the transition state). alphaD97 mutations in the two alpha-subunits have unequal energetic consequences for gating, but their contributions are independent. We conclude that the key, underlying functional consequence of alphaD97 perturbations is to increase the unliganded gating equilibrium constant. L5 emerges as an important and early link in the AChR gating reaction which, in the absence of agonist, serves to increase the relative stability of the closed conformation of the protein.
Figures