

A subset of membrane-associated proteins is ubiquitinated in response to mutations in the endoplasmic reticulum degradation machinery
- PMID: 14557538
- PMCID: PMC240687
- DOI: 10.1073/pnas.2135500100
A subset of membrane-associated proteins is ubiquitinated in response to mutations in the endoplasmic reticulum degradation machinery
Abstract
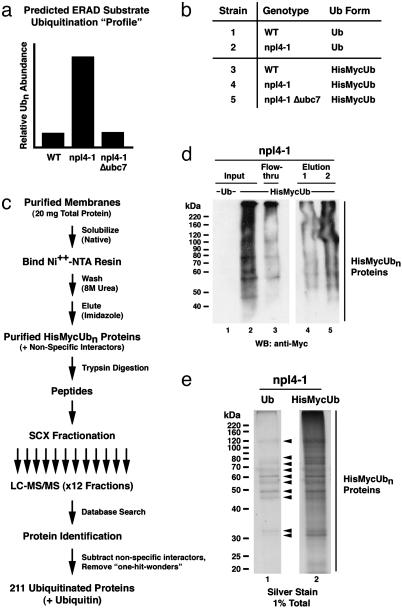
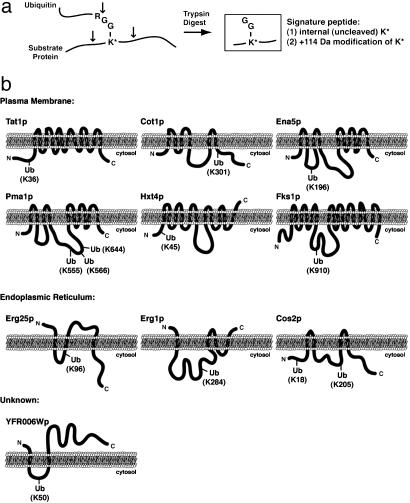
Ubiquitination of membrane-associated proteins can direct their proteasome-mediated degradation or activation at the endoplasmic reticulum (ER), as well as their endocytosis and intracellular sorting. However, the full spectrum of ubiquitinated membrane proteins has not been determined. Here we combined proteomic analysis with yeast genetics to identify 211 ubiquitinated membrane-associated proteins in Saccharomyces cerevisiae and map >30 precise sites of ubiquitination. Major classes of identified ubiquitinated proteins include ER-resident membrane proteins, plasma membrane-localized permeases, receptors, and enzymes, and surprisingly, components of the actin cytoskeleton. By determining the differential abundance of ubiquitinated proteins in yeast mutated for NPL4 and UBC7, which are major components of ER-associated degradation (ERAD), we furthermore were able to classify 83 of these identified ubiquitinated membrane proteins as potential endogenous substrates of the ERAD pathway. These substrates are highly enriched for proteins that localize to or transit through the ER. Interestingly, we also identified novel membrane-bound transcription factors that may be subject to ubiquitin/proteasome-mediated cleavage and activation at the ER membrane.
Figures


References
-
- Plemper, R. K. & Wolf, D. H. (1999) Trends Biochem. Sci. 24 266–270. - PubMed
-
- Deak, P. M. & Wolf, D. H. (2001) J. Biol. Chem. 276 10663–10669. - PubMed
-
- Biederer, T., Volkwein, C. & Sommer, T. (1997) Science 278 1806–1809. - PubMed
-
- Bays, N. W., Gardner, R. G., Seelig, L. P., Joazeiro, C. A. & Hampton, R. Y. (2001) Nat. Cell Biol. 3 24–29. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
