

Role of the serotonin 5-HT(2A) receptor in learning
- PMID: 14557608
- PMCID: PMC218001
- DOI: 10.1101/lm.60803
Role of the serotonin 5-HT(2A) receptor in learning
Abstract
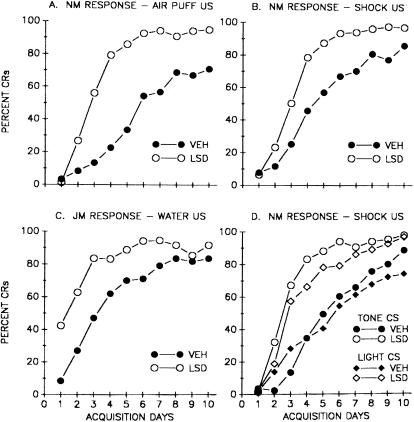
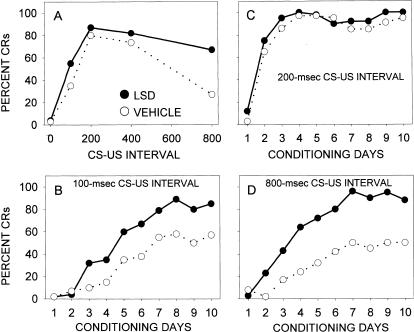
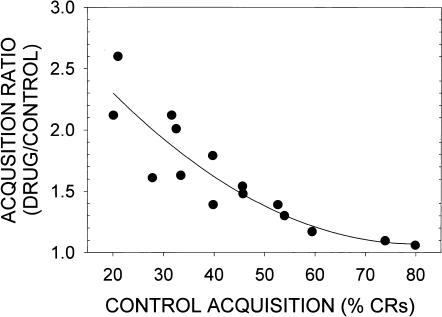
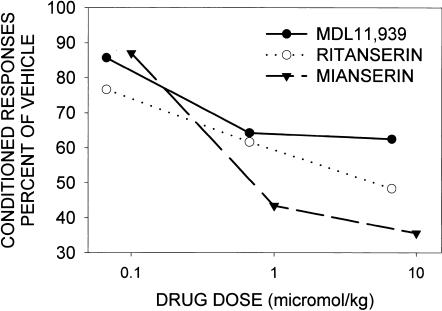
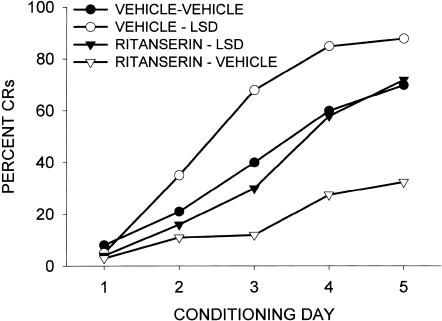
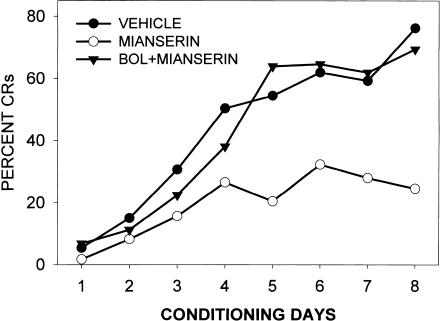
This study reviews the role of the serotonin 5-HT2A receptor in learning as measured by the acquisition of the rabbit's classically conditioning nictitating membrane response, a component of the eyeblink response. Agonists at the 5-HT2A receptor including LSD (d-lysergic acid diethylamide) enhanced associative learning at doses that produce cognitive effects in humans. Some antagonists such as BOL (d-bromolysergic acid diethylamide), LY53,857, and ketanserin acted as neutral antagonists in that they had no effect on learning, whereas others (MDL11,939, ritanserin, and mianserin) acted as inverse agonists in that they retarded learning through an action at the 5-HT2A receptor. These results were placed in the context of what is known concerning the anatomical distribution and electrophysiological effects of 5-HT2A receptor activation in frontal cortex and hippocampus, as well as the role of cortical 5-HT2A receptors in schizophrenia. It was concluded that the 5-HT2A receptor demonstrates constitutive activity, and that variations in this activity can produce profound alterations in cognitive states.
Figures
References
-
- Abramson, H.A., Jarvik, M.E., Kaufman, M.R., Kornetsky, C., Levine, A., and Wagner, M. 1955. Lysergic acid diethylamide (LSD-25). I. Physiological and perceptual responses. J. Psychol. 39: 3–60.
-
- Aghajanian, G.K. and Marek, G.J. 1997. Serotonin induces excitatory postsynaptic potentials in apical dendrites of neocortical pyramidal cells. Neuropharmacology 36: 589–599. - PubMed
-
- Aghajanian, G.K. and Marek, G.J. 2000. Serotonin model of schizophrenia: Emerging role of glutamate mechanisms. Brain Res. Brain Res. Rev. 31: 302–312. - PubMed
-
- Alhaider, A.A., Ageel, A.M., and Ginawi, O.T. 1993. The quipazine- and TFMPP-increased conditioned avoidance response in rats: Role of 5-HT1C/5-HT2 receptors. Neuropharmacology 32: 1427–1432. - PubMed
-
- Aloyo, V.J. and Harvey, J.A. 2000. Antagonist binding at 5-HT2A and 5-HT2C receptors in the rabbit: High correlation with the profile for the human receptors. Eur. J. Pharmacol. 406: 163–169. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources