

Measles virus (MV) nucleoprotein binds to a novel cell surface receptor distinct from FcgammaRII via its C-terminal domain: role in MV-induced immunosuppression
- PMID: 14557619
- PMCID: PMC229257
- DOI: 10.1128/jvi.77.21.11332-11346.2003
Measles virus (MV) nucleoprotein binds to a novel cell surface receptor distinct from FcgammaRII via its C-terminal domain: role in MV-induced immunosuppression
Abstract
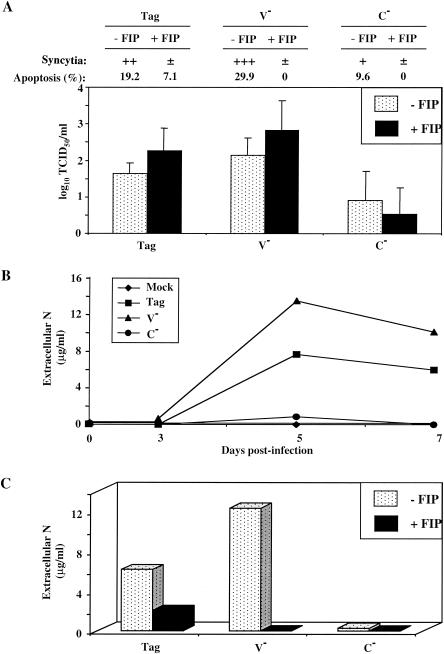
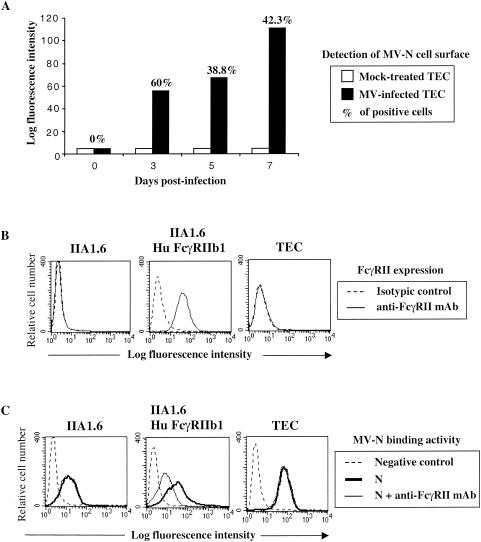
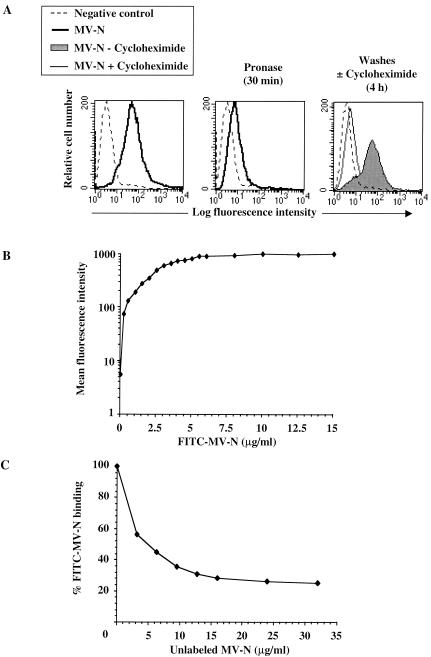
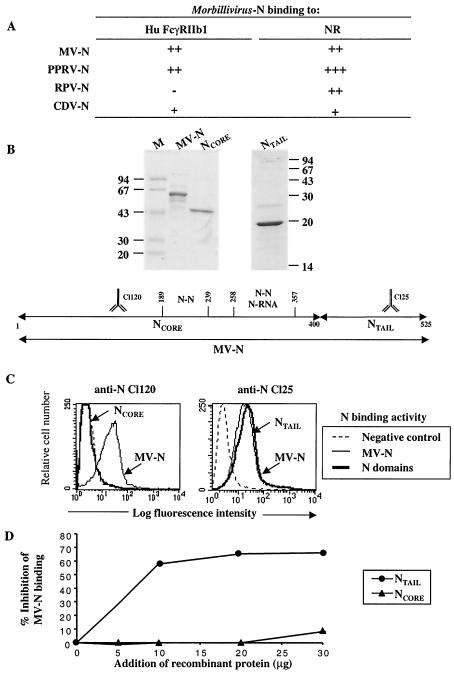
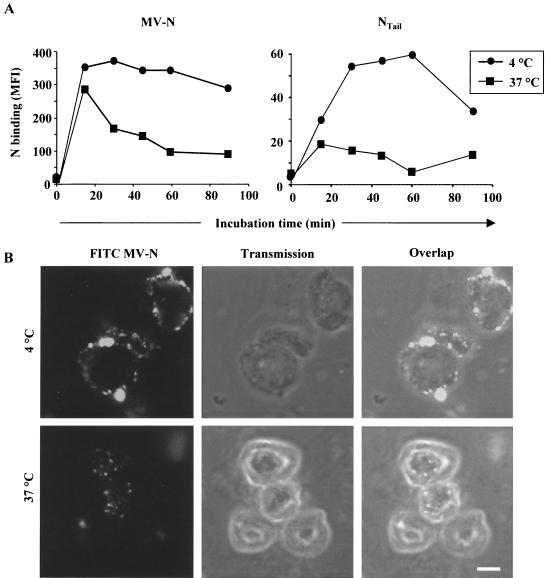
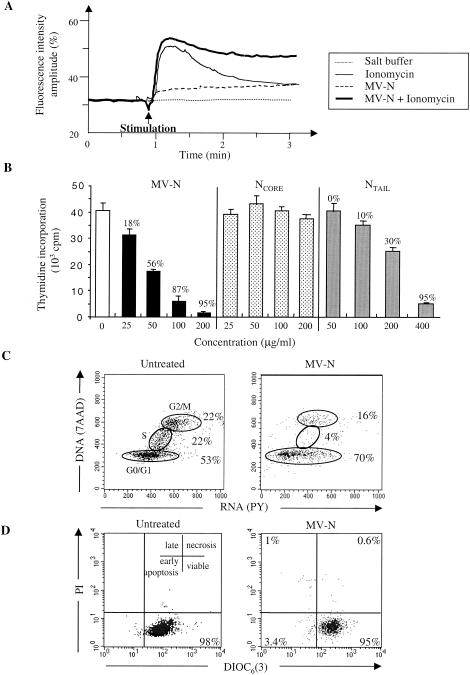
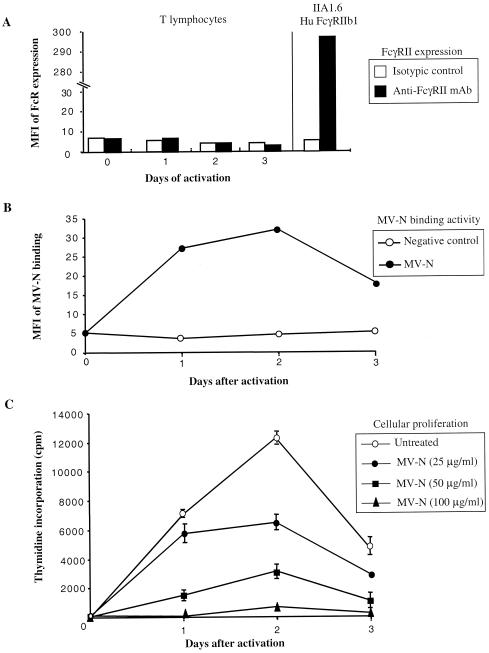
During acute measles virus (MV) infection, an efficient immune response occurs, followed by a transient but profound immunosuppression. MV nucleoprotein (MV-N) has been reported to induce both cellular and humoral immune responses and paradoxically to account for immunosuppression. Thus far, this latter activity has been attributed to MV-N binding to human and murine FcgammaRII. Here, we show that apoptosis of MV-infected human thymic epithelial cells (TEC) allows the release of MV-N in the extracellular compartment. This extracellular N is then able to bind either to MV-infected or uninfected TEC. We show that recombinant MV-N specifically binds to a membrane protein receptor, different from FcgammaRII, highly expressed on the cell surface of TEC. This new receptor is referred to as nucleoprotein receptor (NR). In addition, different Ns from other MV-related morbilliviruses can also bind to FcgammaRII and/or NR. We show that the region of MV-N responsible for binding to NR maps to the C-terminal fragment (N(TAIL)). Binding of MV-N to NR on TEC triggers sustained calcium influx and inhibits spontaneous cell proliferation by arresting cells in the G(0) and G(1) phases of the cell cycle. Finally, MV-N binds to both constitutively expressed NR on a large spectrum of cells from different species and to human activated T cells, leading to suppression of their proliferation. These results provide evidence that MV-N, after release in the extracellular compartment, binds to NR and thereby plays a role in MV-induced immunosuppression.
Figures
Similar articles
-
Measles virus nucleoprotein induces cell-proliferation arrest and apoptosis through NTAIL-NR and NCORE-FcgammaRIIB1 interactions, respectively.J Gen Virol. 2005 Jun;86(Pt 6):1771-1784. doi: 10.1099/vir.0.80791-0. J Gen Virol. 2005. PMID: 15914856
-
Cell surface delivery of the measles virus nucleoprotein: a viral strategy to induce immunosuppression.J Virol. 2004 Nov;78(21):11952-61. doi: 10.1128/JVI.78.21.11952-11961.2004. J Virol. 2004. PMID: 15479835 Free PMC article.
-
Measles virus nucleocapsid protein binds to FcgammaRII and inhibits human B cell antibody production.J Exp Med. 1997 Jul 21;186(2):269-78. doi: 10.1084/jem.186.2.269. J Exp Med. 1997. PMID: 9221756 Free PMC article.
-
Immunosuppression caused by measles virus: role of viral proteins.Rev Med Virol. 2006 Jan-Feb;16(1):49-63. doi: 10.1002/rmv.486. Rev Med Virol. 2006. PMID: 16237742 Review.
-
Nectin 4 is the epithelial cell receptor for measles virus.Trends Microbiol. 2012 Sep;20(9):429-39. doi: 10.1016/j.tim.2012.05.006. Epub 2012 Jun 20. Trends Microbiol. 2012. PMID: 22721863 Review.
Cited by
-
Measles virus-induced immunosuppression: from effectors to mechanisms.Med Microbiol Immunol. 2010 Aug;199(3):227-37. doi: 10.1007/s00430-010-0152-3. Epub 2010 Apr 8. Med Microbiol Immunol. 2010. PMID: 20376484 Review.
-
The intrinsically disordered C-terminal domain of the measles virus nucleoprotein interacts with the C-terminal domain of the phosphoprotein via two distinct sites and remains predominantly unfolded.Protein Sci. 2005 Aug;14(8):1975-92. doi: 10.1110/ps.051411805. Protein Sci. 2005. PMID: 16046624 Free PMC article.
-
The measles virus nucleocapsid protein tail domain is dispensable for viral polymerase recruitment and activity.J Biol Chem. 2013 Oct 11;288(41):29943-53. doi: 10.1074/jbc.M113.503862. Epub 2013 Sep 3. J Biol Chem. 2013. PMID: 24003217 Free PMC article.
-
Cell surface RNA virus nucleocapsid proteins: a viral strategy for immunosuppression?Npj Viruses. 2024 Sep 2;2(1):41. doi: 10.1038/s44298-024-00051-3. Npj Viruses. 2024. PMID: 40295865 Free PMC article. Review.
-
The thymus is a common target organ in infectious diseases.PLoS Pathog. 2006 Jun;2(6):e62. doi: 10.1371/journal.ppat.0020062. PLoS Pathog. 2006. PMID: 16846255 Free PMC article. Review.
References
-
- Bankamp, B., S. M. Horikami, P. D. Thompson, M. Huber, M. Billeter, and S. A. Moyer. 1996. Domains of the measles virus N protein required for binding to P protein and self-assembly. Virology 216:272-277. - PubMed
-
- Barrett, T. 1999. Morbillivirus infections, with special emphasis on morbilliviruses of carnivores. Vet. Microbiol. 69:3-13. - PubMed
-
- Bhella, D., A. Ralph, L. B. Murphy, and R. P. Yeo. 2002. Significant differences in nucleocapsid morphology within the Paramyxoviridae. J. Gen. Virol. 83:1831-1839. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
