

PU.1 and pRB interact and cooperate to repress GATA-1 and block erythroid differentiation
- PMID: 14559995
- PMCID: PMC207581
- DOI: 10.1128/MCB.23.21.7460-7474.2003
PU.1 and pRB interact and cooperate to repress GATA-1 and block erythroid differentiation
Abstract
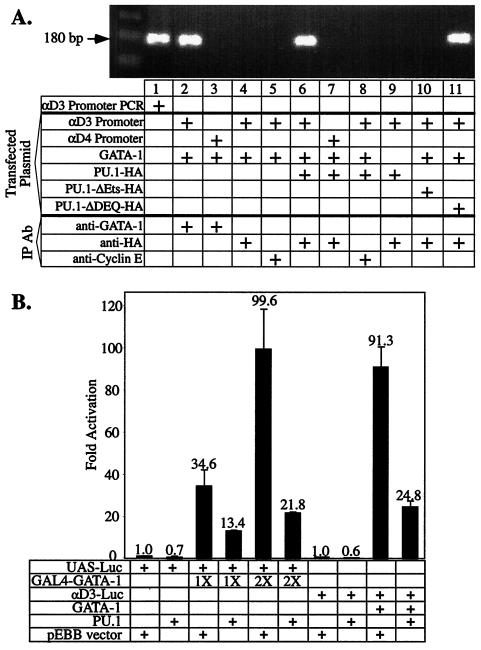
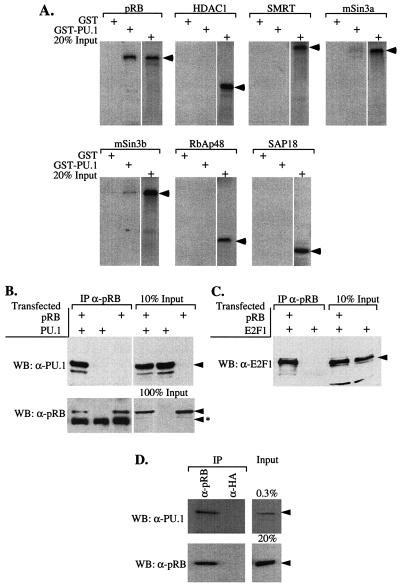
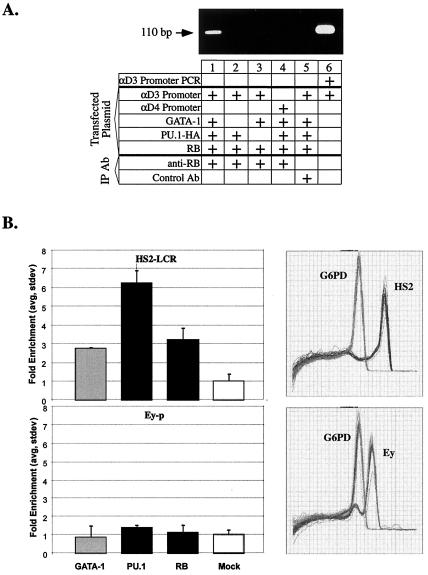
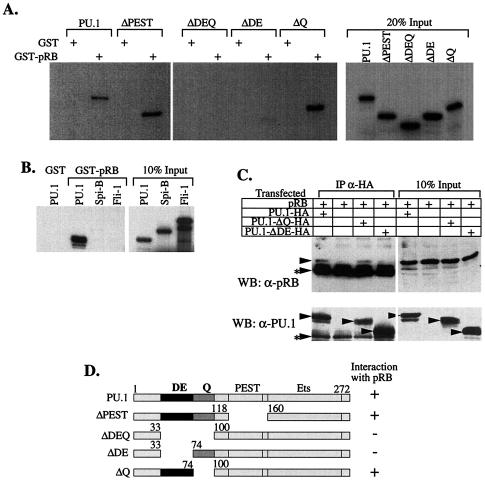
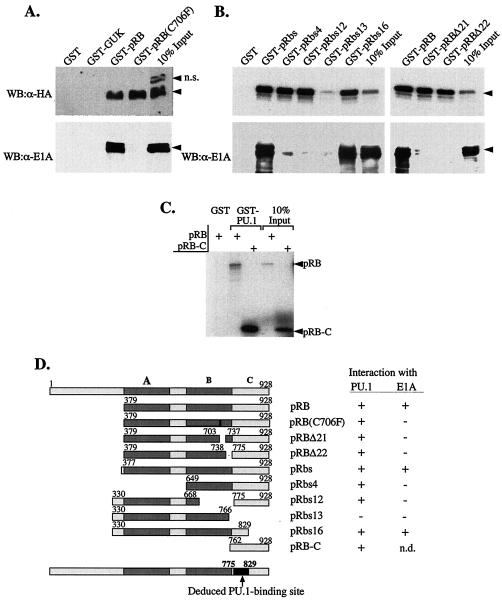
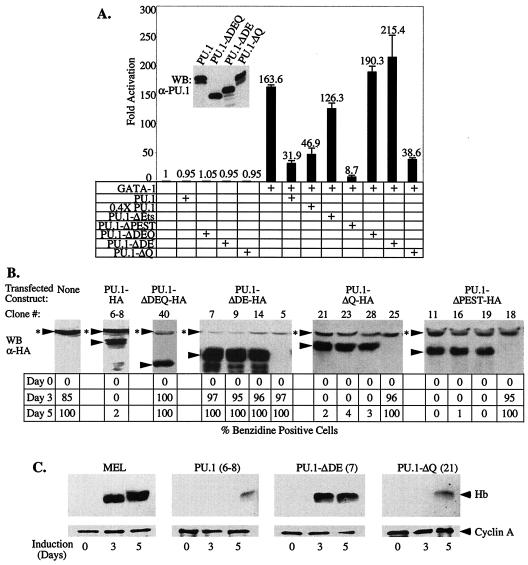
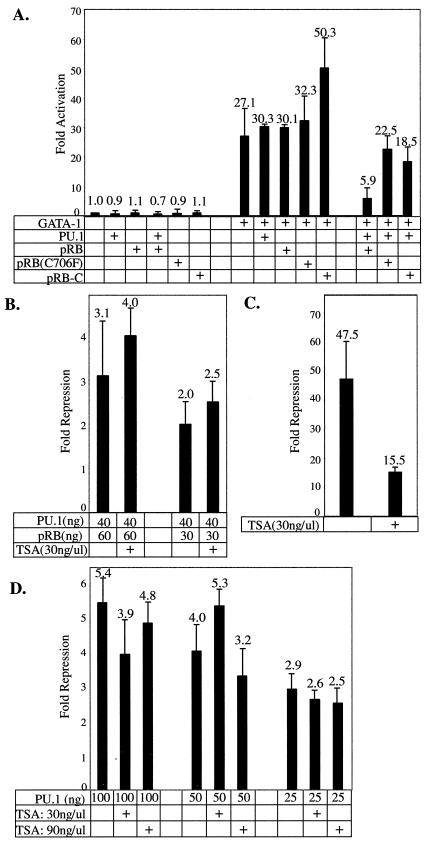
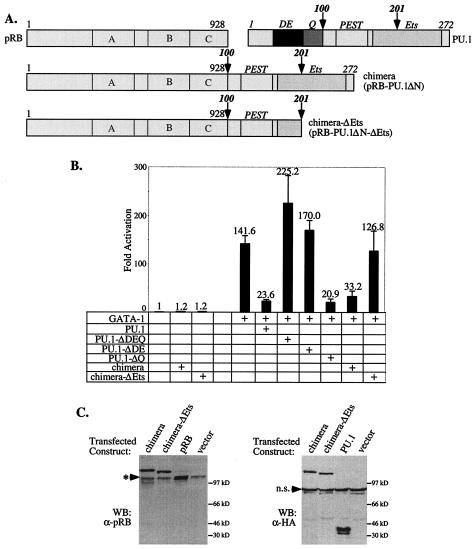
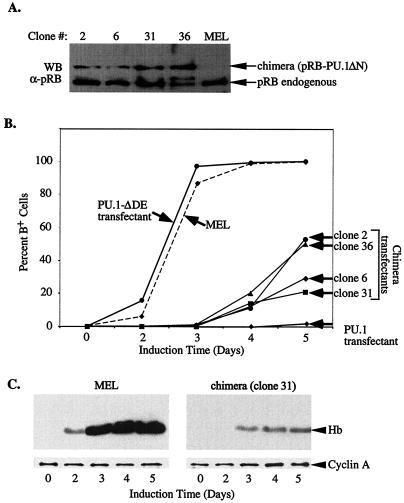
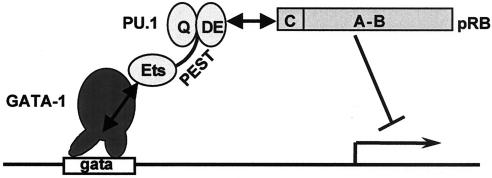
PU.1 and GATA-1 are two hematopoietic specific transcription factors that play key roles in development of the myeloid and erythroid lineages, respectively. The two proteins bind to one another and inhibit each other's function in transcriptional activation and promotion of their respective differentiation programs. This mutual antagonism may be an important aspect of lineage commitment decisions. PU.1 can also act as an oncoprotein since deregulated expression of PU.1 in erythroid precursors causes erythroleukemias in mice. Studies of cultured mouse erythroleukemia cell lines indicate that one aspect of PU.1 function in erythroleukemogenesis is its ability to block erythroid differentiation by repressing GATA-1 (N. Rekhtman, F. Radparvar, T. Evans, and A. I. Skoultchi, Genes Dev. 13:1398-1411, 1999). We have investigated the mechanism of PU.1-mediated repression of GATA-1. We report here that PU.1 binds to GATA-1 on DNA. We localized the repression activity of PU.1 to a small acidic N-terminal domain that interacts with the C pocket of pRB, a well-known transcriptional corepressor. Repression of GATA-1 by PU.1 requires pRB, and pRB colocalizes with PU.1 and GATA-1 at repressed GATA-1 target genes. PU.1 and pRB also cooperate to block erythroid differentiation. Our results suggest that one of the mechanisms by which PU.1 antagonizes GATA-1 is by binding to it at GATA-1 target genes and tethering to these sites a corepressor that blocks transcriptional activity and thereby erythroid differentiation.
Figures
References
-
- Aumont, F. L., P. Trudel, and L. Wall. 1993. Murine erythroleukemia cells contain two distinct GATA-binding proteins that have different patterns of expression during cellular differentiation. Differentiation 52:169-176. - PubMed
-
- Behre, G., A. J. Whitmarsh, M. P. Coghlan, T. Hoang, C. L. Carpenter, D. E. Zhang, R. J. Davis, and D. G. Tenen. 1999. c-Jun is a JNK-independent coactivator of the PU.1 transcription factor. J. Biol. Chem. 274:4939-4946. - PubMed
-
- Ben-David, Y., and A. Bernstein. 1991. Friend virus-induced erythroleukemia and the multistage nature of cancer. Cell 66:831-834. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources