

Elimination of protein kinase MK5/PRAK activity by targeted homologous recombination
- PMID: 14560018
- PMCID: PMC207624
- DOI: 10.1128/MCB.23.21.7732-7741.2003
Elimination of protein kinase MK5/PRAK activity by targeted homologous recombination
Abstract
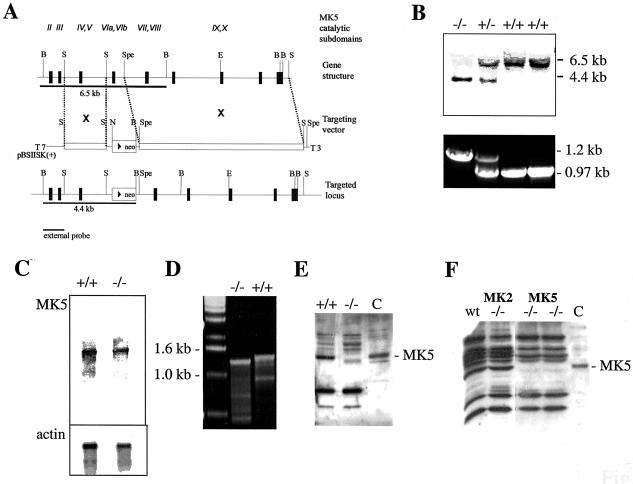
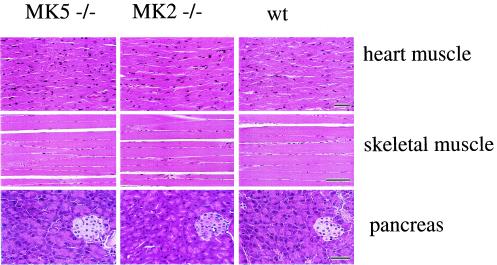
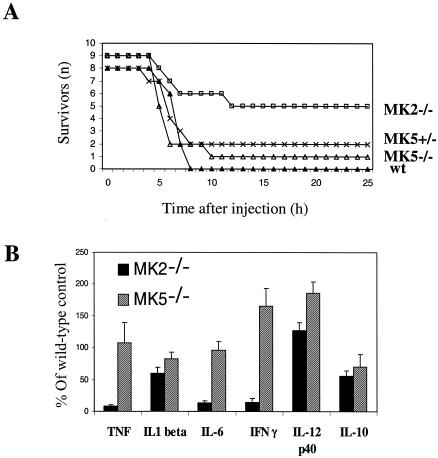
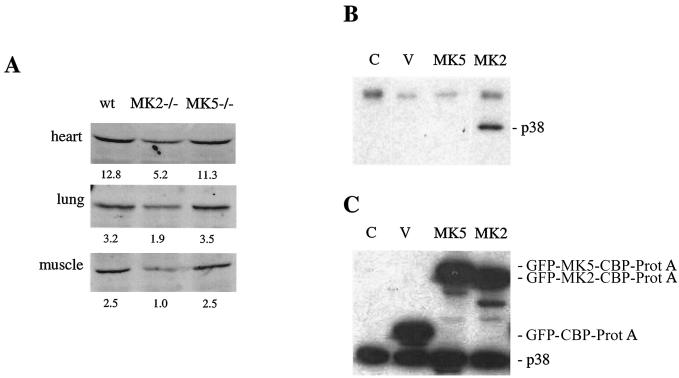
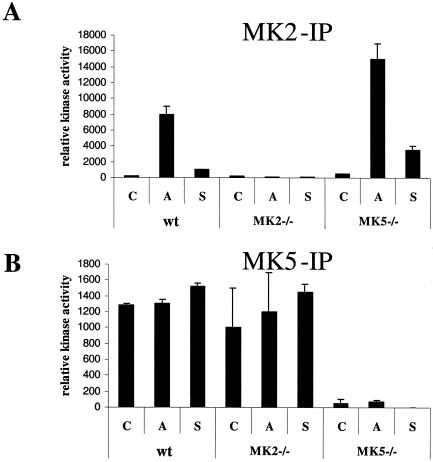
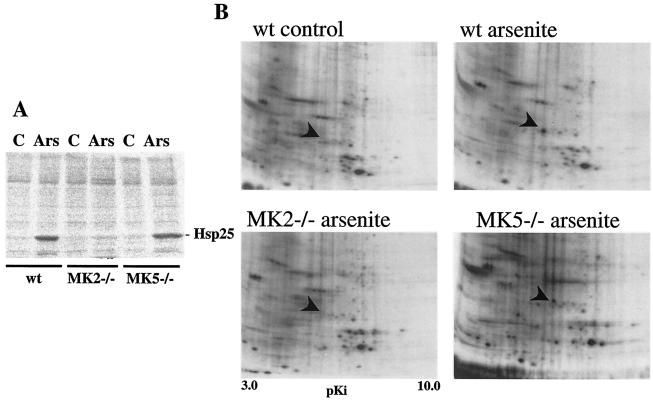
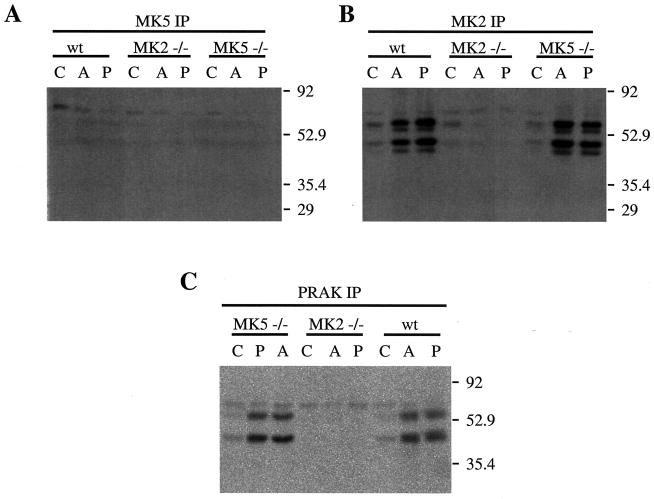
MK5 (mitogen-activated protein kinase [MAPK]-activated protein kinase 5), also designated PRAK (p38-regulated and -activated kinase), was deleted from mice by homologous recombination. Although no MK5 full-length protein and kinase activity was detected in the MK5 knockout mice, the animals were viable and fertile and did not display abnormalities in tissue morphology or behavior. In addition, these mice did not show increased resistance to endotoxic shock or decreased lipopolysaccharide-induced cytokine production. Hence, MK5 deletion resulted in a phenotype very different from the complex inflammation-impaired phenotype of mice deficient in MK2, although MK2 and MK5 exhibit evolutional, structural, and apparent extensive functional similarities. To explain this discrepancy, we used wild-type cells and embryonic fibroblasts from both MK2 and MK5 knockout mice as controls to reexamine the mechanism of activation, the interaction with endogenous p38 MAPK, and the substrate specificity of both enzymes. In contrast to MK2, which shows interaction with and chaperoning properties for p38 MAPK and which is activated by extracellular stresses such as arsenite or sorbitol treatment, endogenous MK5 did not show these properties. Furthermore, endogenous MK5 is not able to phosphorylate Hsp27 in vitro and in vivo. We conclude that the differences between the phenotypes of MK5- and MK2-deficient mice result from clearly different functional properties of both enzymes.
Figures
Similar articles
-
Does MK5 reconcile classical and atypical MAP kinases?Front Biosci. 2008 May 1;13:4617-24. doi: 10.2741/3027. Front Biosci. 2008. PMID: 18508533
-
Both binding and activation of p38 mitogen-activated protein kinase (MAPK) play essential roles in regulation of the nucleocytoplasmic distribution of MAPK-activated protein kinase 5 by cellular stress.Mol Cell Biol. 2002 Oct;22(20):6931-45. doi: 10.1128/MCB.22.20.6931-6945.2002. Mol Cell Biol. 2002. PMID: 12242275 Free PMC article.
-
Distinct cellular functions of MK2.Mol Cell Biol. 2002 Jul;22(13):4827-35. doi: 10.1128/MCB.22.13.4827-4835.2002. Mol Cell Biol. 2002. PMID: 12052889 Free PMC article.
-
Mitogen-activated protein kinase p38 and MK2, MK3 and MK5: ménage à trois or ménage à quatre?Cell Signal. 2010 Aug;22(8):1185-92. doi: 10.1016/j.cellsig.2010.03.002. Epub 2010 Mar 11. Cell Signal. 2010. PMID: 20227494 Review.
-
MK5: A novel regulator of cardiac fibroblast function?IUBMB Life. 2017 Oct;69(10):785-794. doi: 10.1002/iub.1677. Epub 2017 Sep 11. IUBMB Life. 2017. PMID: 28941148 Review.
Cited by
-
ERK and p38 MAPK-activated protein kinases: a family of protein kinases with diverse biological functions.Microbiol Mol Biol Rev. 2004 Jun;68(2):320-44. doi: 10.1128/MMBR.68.2.320-344.2004. Microbiol Mol Biol Rev. 2004. PMID: 15187187 Free PMC article. Review.
-
Comparative molecular dynamics simulations of mitogen-activated protein kinase-activated protein kinase 5.Int J Mol Sci. 2014 Mar 19;15(3):4878-902. doi: 10.3390/ijms15034878. Int J Mol Sci. 2014. PMID: 24651460 Free PMC article.
-
Characterization of hsp27 kinases activated by elevated aortic pressure in heart.Mol Cell Biochem. 2012 Dec;371(1-2):31-42. doi: 10.1007/s11010-012-1420-x. Epub 2012 Aug 10. Mol Cell Biochem. 2012. PMID: 22878564 Free PMC article.
-
Tumour promoting and suppressing roles of the atypical MAP kinase signalling pathway ERK3/4-MK5.J Mol Signal. 2012 Jul 16;7(1):9. doi: 10.1186/1750-2187-7-9. J Mol Signal. 2012. PMID: 22800433 Free PMC article.
-
Serine residue 115 of MAPK-activated protein kinase MK5 is crucial for its PKA-regulated nuclear export and biological function.Cell Mol Life Sci. 2011 Mar;68(5):847-62. doi: 10.1007/s00018-010-0496-2. Epub 2010 Aug 25. Cell Mol Life Sci. 2011. PMID: 20734105 Free PMC article.
References
-
- Ben-Levy, R., S. Hooper, R. Wilson, H. F. Paterson, and C. J. Marshall. 1998. Nuclear export of the stress-activated protein kinase p38 mediated by its substrate MAPKAP kinase-2. Curr. Biol. 8:1049-1057. - PubMed
-
- Clifton, A. D., P. R. Young, and P. Cohen. 1996. A comparison of the substrate specificity of MAPKAP kinase-2 and MAPKAP kinase-3 and their activation by cytokines and cellular stress. FEBS Lett. 392:209-214. - PubMed
-
- Engel, K., K. Plath, and M. Gaestel. 1993. The MAP kinase-activated protein kinase 2 contains a proline-rich SH3-binding domain. FEBS Lett. 336:143-147. - PubMed
-
- Engel, K., H. Schultz, F. Martin, A. Kotlyarov, K. Plath, M. Hahn, U. Heinemann, and M. Gaestel. 1995. Constitutive activation of mitogen-activated protein kinase-activated protein kinase 2 by mutation of phosphorylation sites and an A-helix motif. J. Biol. Chem. 270:27213-27221. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous