

Mutations in yeast replication proteins that increase CAG/CTG expansions also increase repeat fragility
- PMID: 14560028
- PMCID: PMC207578
- DOI: 10.1128/MCB.23.21.7849-7860.2003
Mutations in yeast replication proteins that increase CAG/CTG expansions also increase repeat fragility
Abstract
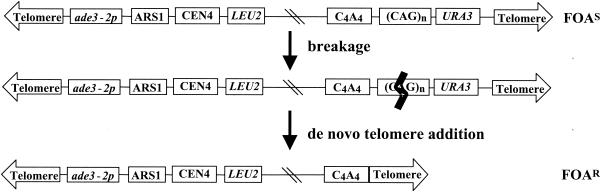
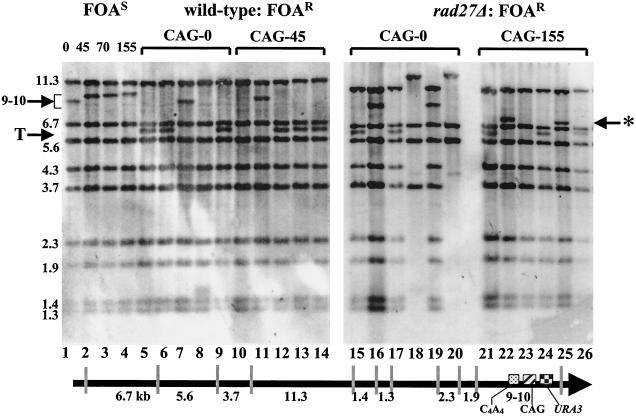
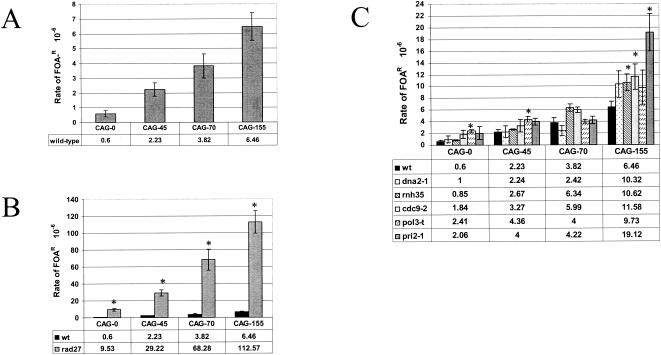
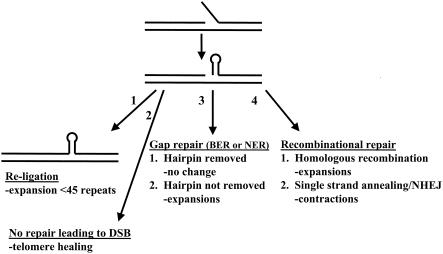
Expansion of trinucleotide repeats (TNRs) is the causative mutation in several human genetic diseases. Expanded TNR tracts are both unstable (changing in length) and fragile (displaying an increased propensity to break). We have investigated the relationship between fidelity of lagging-strand replication and both stability and fragility of TNRs. We devised a new yeast artificial chromomosme (YAC)-based assay for chromosome breakage to analyze fragility of CAG/CTG tracts in mutants deficient for proteins involved in lagging-strand replication: Fen1/Rad27, an endo/exonuclease involved in Okazaki fragment maturation, the nuclease/helicase Dna2, RNase HI, DNA ligase, polymerase delta, and primase. We found that deletion of RAD27 caused a large increase in breakage of short and long CAG/CTG tracts, and defects in DNA ligase and primase increased breakage of long tracts. We also found a correlation between mutations that increase CAG/CTG tract breakage and those that increase repeat expansion. These results suggest that processes that generate strand breaks, such as faulty Okazaki fragment processing or DNA repair, are an important source of TNR expansions.
Figures
References
-
- Anderson, S., and M. L. DePamphilis. 1979. Metabolism of Okazaki fragments during simian virus 40 DNA replication. J. Biol. Chem. 254:11495-11504. - PubMed
-
- Ayyagari, R., X. V. Gomes, D. A. Gordenin, and P. M. Burgers. 2003. Okazaki fragment maturation in yeast. I. Distribution of functions between FEN1 and DNA2. J. Biol. Chem. 278:1618-1625. - PubMed
-
- Bae, S. H., K. H. Bae, J. A. Kim, and Y. S. Seo. 2001. RPA governs endonuclease switching during processing of Okazaki fragments in eukaryotes. Nature 412:456-461. - PubMed
-
- Bae, S.-H., E. Choi, K.-H. Lee, J. S. Park, S.-H. Lee, and Y.-S. Seo. 1998. Dna2 of Saccharomyces cerevisiae possesses a single-stranded DNA-specific endonuclease activity that is able to act on double-stranded DNA in the presence of ATP. J. Biol. Chem. 273:26880-26890. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous