

Interactions between centromere complexes in Saccharomyces cerevisiae
- PMID: 14565975
- PMCID: PMC284796
- DOI: 10.1091/mbc.e03-06-0419
Interactions between centromere complexes in Saccharomyces cerevisiae
Abstract
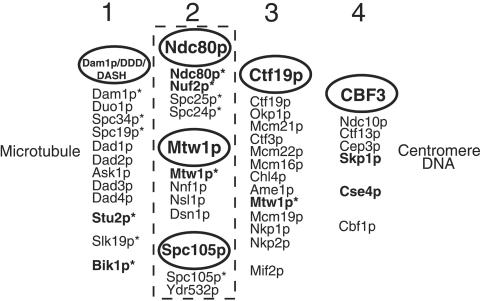
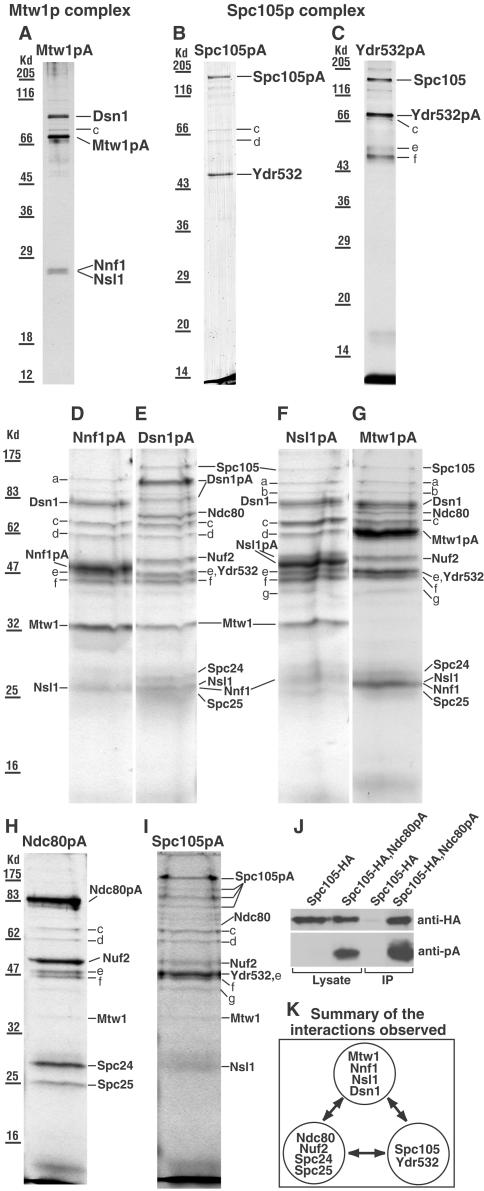
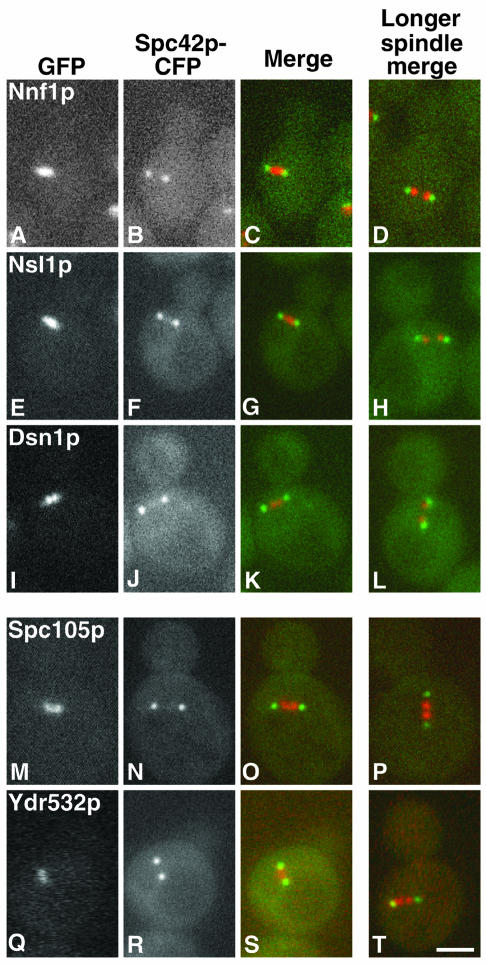
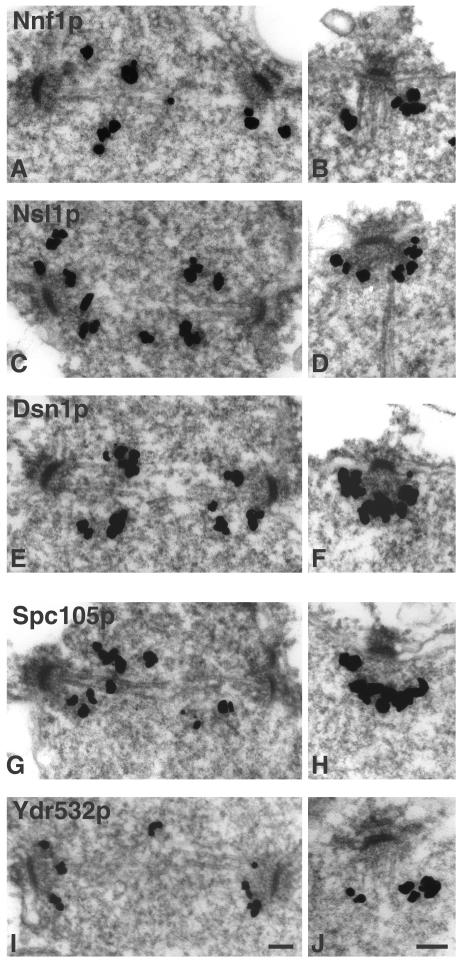
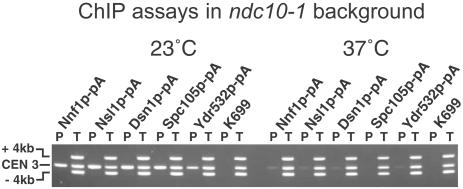
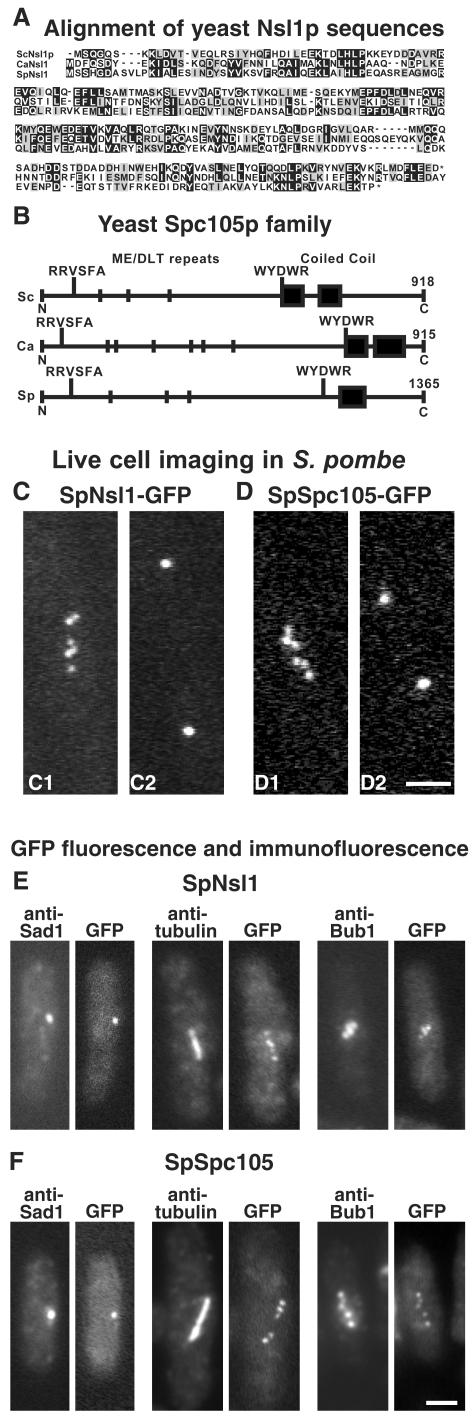
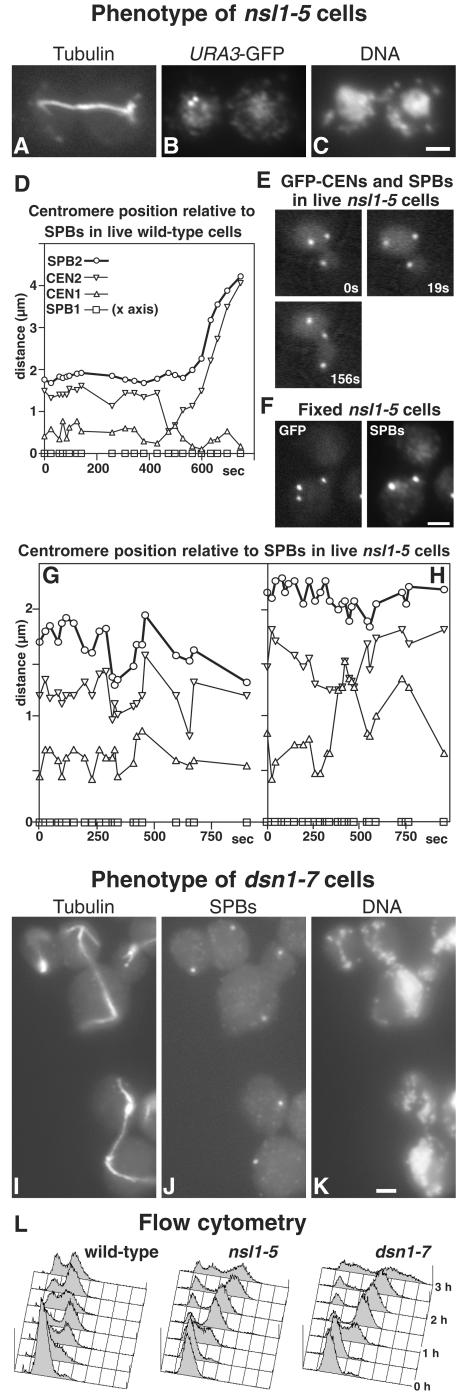
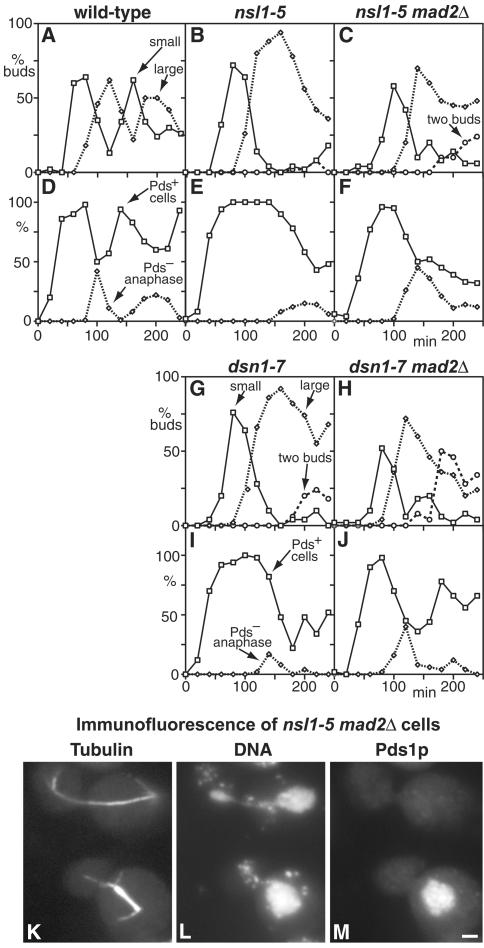
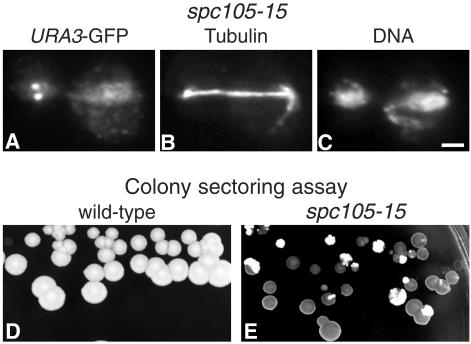
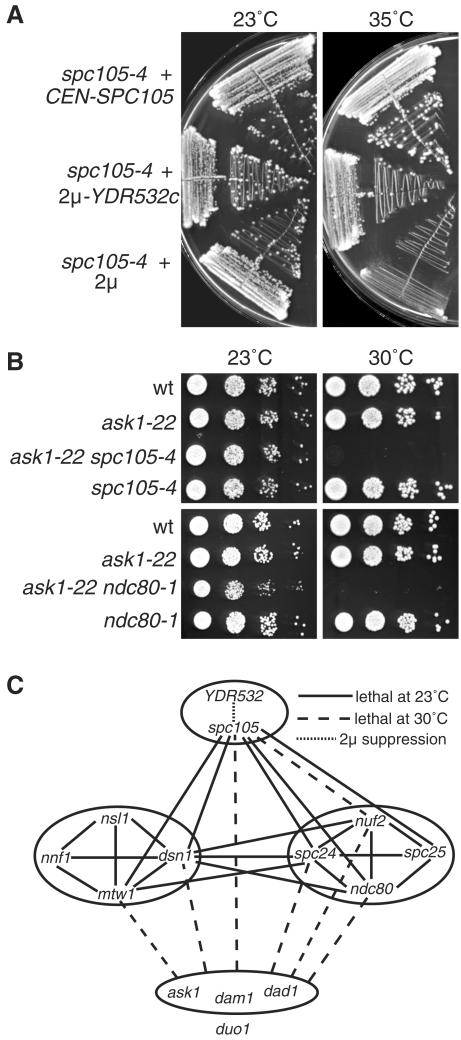
We have purified two new complexes from Saccharomyces cerevisiae, one containing the centromere component Mtw1p together with Nnf1p, Nsl1p, and Dsn1p, which we call the Mtw1p complex, and the other containing Spc105p and Ydr532p, which we call the Spc105p complex. Further purifications using Dsn1p tagged with protein A show, in addition to the other components of the Mtw1p complex, the two components of the Spc105p complex and the four components of the previously described Ndc80p complex, suggesting that all three complexes are closely associated. Fluorescence microscopy and immunoelectron microscopy show that Nnf1p, Nsl1p, Dsn1p, Spc105p, and Ydr532p all localize to the nuclear side of the spindle pole body and along short spindles. Chromatin immunoprecipitation assays show that all five proteins are associated with centromere DNA. Homologues of Nsl1p and Spc105p in Schizosaccharomyces pombe also localize to the centromere. Temperature-sensitive mutations of Nsl1p, Dsn1p, and Spc105p all cause defects in chromosome segregation. Synthetic-lethal interactions are found between temperature-sensitive mutations in proteins from all three complexes, in agreement with their close physical association. These results show an increasingly complex structure for the S. cerevisiae centromere and a probable conservation of structure between parts of the centromeres of S. cerevisiae and S. pombe.
Figures
References
-
- Aebersold, R., and Mann, M. (2003). Mass spectrometry-based proteomics. Nature 422, 198–207. - PubMed
-
- Bähler, J., Wu, J.-Q., Longtine, M.S., Shah, N.G., McKenzie, A., III, Steever, A.B., Wach, A., Philippsen, P., and Pringle, J.R. (1998). Heterologous modules for efficient and versatile PCR-based gene targeting in Schizosaccharomyces pombe. Yeast 14, 943–951. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
