

Cell-specific expression of CYP2A5 in the mouse respiratory tract: effects of olfactory toxicants
- PMID: 14566026
- PMCID: PMC3957563
- DOI: 10.1177/002215540305101114
Cell-specific expression of CYP2A5 in the mouse respiratory tract: effects of olfactory toxicants
Abstract
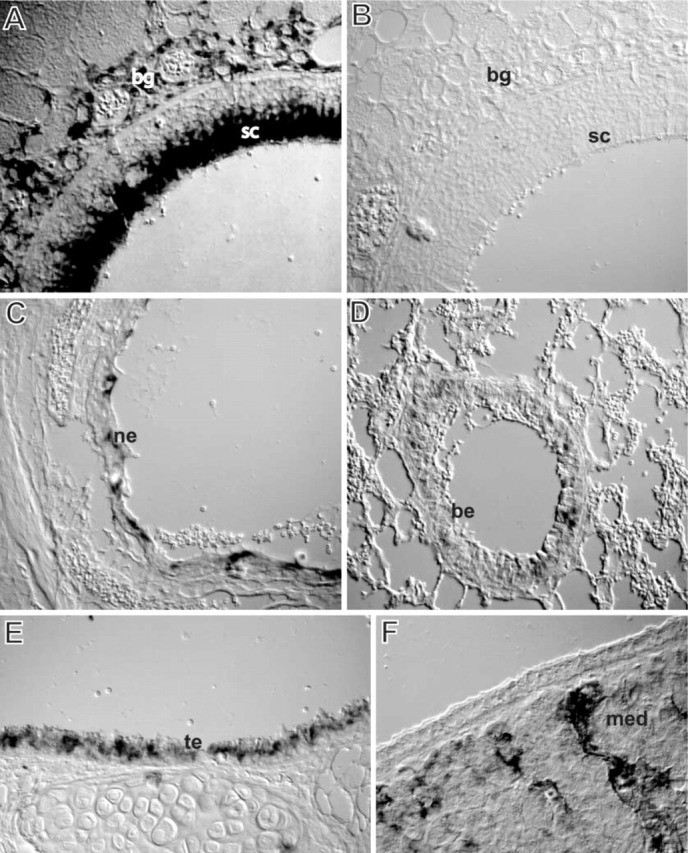
We performed a detailed analysis of mouse cytochrome P450 2A5 (CYP2A5) expression by in situ hybridization (ISH) and immunohistochemistry (IHC) in the respiratory tissues of mice. The CYP2A5 mRNA and the corresponding protein co-localized at most sites and were predominantly detected in the olfactory region, with an expression in sustentacular cells, Bowman's gland, and duct cells. In the respiratory and transitional epithelium there was no or only weak expression. The nasolacrimal duct and the excretory ducts of nasal and salivary glands displayed expression, whereas no expression occurred in the acini. There was decreasing expression along the epithelial linings of the trachea and lower respiratory tract, whereas no expression occurred in the alveoli. The hepatic CYP2A5 inducers pyrazole and phenobarbital neither changed the CYP2A5 expression pattern nor damaged the olfactory mucosa. In contrast, the olfactory toxicants dichlobenil and methimazole induced characteristic changes. The damaged Bowman's glands displayed no expression, whereas the damaged epithelium expressed the enzyme. The CYP2A5 expression pattern is in accordance with previously reported localization of protein and DNA adducts and the toxicity of some CYP2A5 substrates. This suggests that CYP2A5 is an important determinant for the susceptibility of the nasal and respiratory epithelia to protoxicants and procarcinogens.
Figures


References
-
- Adams DR, Jones AM, Plopper CG, Serabjit—Singh CJ, Philpot RM. (1991) Distribution of cytochrome P-450 monoxygenase enzymes in the nasal mucosa of hamster and rat. Am J Anat 190:291–298 - PubMed
-
- Bakke JE, Larsen GL, Struble C, Feil VJ, Brandt I, Brittebo EB. (1988) Metabolism of 2,6-dichlorobenzonitrile, 2,6-dichlorothiobenzamide in rodents and goats. Xenobiotica 18:1063–1075 - PubMed
-
- Belinsky SA, Foley JF, White CM, Anderson MW, Maronpot RR. (1990) Dose-response relationship between O6-methylguanine formation in Clara cells and induction of pulmonary neoplasia in the rat by 4-(methylnitrosamino)-1-(3-pyridyl)-1-butanone. Cancer Res 50:3772–3780 - PubMed
-
- Belinsky SA, Walker VE, Maronpot RR, Swenberg JA, Anderson MW. (1987) Molecular dosimetry of DNA adduct formation and cell toxicity in rat nasal mucosa following exposure to the tobacco specific nitrosamine 4-(N-methyl-N-nitrosamino)-1-(3-pyridyl)-1-butanone and their relationship to induction of neoplasia. Cancer Res 47:6058–6065 - PubMed
-
- Bereziat JC, Raffalli F, Schmezer P, Frei E, Geneste O, Lang MA. (1995) Cytochrome P450 2A of nasal epithelium: regulation and role in carcinogen metabolism. Mol Carcinogen 14:130–139 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
