

doi: 10.1016/s0092-8674(03)00803-1.
Nitric oxide regulates exocytosis by S-nitrosylation of N-ethylmaleimide-sensitive factor
Affiliations
- PMID: 14567912
- PMCID: PMC2846406
- DOI: 10.1016/s0092-8674(03)00803-1
Item in Clipboard
Nitric oxide regulates exocytosis by S-nitrosylation of N-ethylmaleimide-sensitive factor
Cell.
.
Abstract
Nitric oxide (NO) inhibits vascular inflammation, but the molecular basis for its anti-inflammatory properties is unknown. We show that NO inhibits exocytosis of Weibel-Palade bodies, endothelial granules that mediate vascular inflammation and thrombosis, by regulating the activity of N-ethylmaleimide-sensitive factor (NSF). NO inhibits NSF disassembly of soluble NSF attachment protein receptor (SNARE) complexes by nitrosylating critical cysteine residues of NSF. NO may regulate exocytosis in a variety of physiological processes, including vascular inflammation, neurotransmission, thrombosis, and cytotoxic T lymphocyte cell killing.
Figures

(A) Time course. HAEC were pretreated with DEA as a control or the NO donor DEA-NONOate or the NOS inhibitor L-NAME, and then incubated with thrombin for 1 hr. The amount of vWF released from cells into the media was measured by an ELISA (n = 3–5 ± SD). Exogenous NO inhibits exocytosis, and L-NAME inhibition of endogenous NOS increases exocytosis. (B) Dose response. HAEC were pretreated with DEA-NONOate for 10 min, DETA-NONOate for 16 hr, SNAP for 6 hr, or NEM for 1 hr and then incubated with thrombin for 1 hr. The amount of vWF released from cells was measured by an ELISA (n = 3–5 ± SD). (C) Inhibition of endogenous NOS increases vWF release. HAEC were pretreated with L-NAME for 16 hr, incubated with thrombin for 1 hr, and vWF released from cells was measured as above (n = 2 ± SD, *p < 0.06 versus thrombin; **p = 0.03 versus thrombin). (D) Activation of endogenous NOS decreases vWF release. HAEC were pretreated with media or 1 mM L-NAME for 16 hr, stimulated with 50 ng/ml VEGF for 2 hr, and then incubated with thrombin. The amount of released vWF was measured as above (n = 3 ± SD, *p < 0.01 versus control).

(A) NSF and SNARE molecules are expressed in endothelial cells. Immunoblotting of 200 μg extracts of endothelial cells or human brain with antibodies to NSF and SNARE molecules. (B) Antibodies to SNARE molecules inhibit Weibel-Palade body exocytosis. HAEC were permeabilized and incubated with antibodies to SNARE molecules. Cells were then resealed, treated with thrombin, and the amount of vWF released into the media was measured by ELISA (n = 2). (C) Antibody to NSF inhibits Weibel-Palade body exocytosis. HAEC were permeabilized and incubated with antibody to NSF or α-SNAP. Cells were then resealed, treated with thrombin, and the amount of vWF released was measured as above (n = 2). (D) NSF peptides enter HAEC. Cells were incubated with FITC-labeled TAT-NSF peptide for 20 min, treated with ethidium bromide to quench extracellular FITC, and imaged by FACS (n = 3 ± SD). (E) NSF peptides inhibit vWF release. Cells were incubated with TAT-NSF peptides for 20 min, treated with media (left) or thrombin (middle and right), and the amount of released vWF was measured (n = 3 ± SD).

(A) NO does not inhibit NSF ATPase activity. A colorimetric assay was used to measure the ATPase activity of recombinant NSF that had been treated with DEA-NONOate (n = 3 ± SD). (B) NO inhibits NSF disassembly of 20S complex. Recombinant NSF was pretreated or not with 1 mM DEA-NONOate for 10 min, mixed with α-SNAP, and added to detergent extracts of HAEC membranes. ATP or ATP-γS 0.5 mM was added, and the mixture was fractionated by ultracentrifugation. Fractions were collected from the bottom of the tubes and immunoblotted with antibody to syn-taxin-4. (C) NO inhibits NSF disassembly activity. Recombinant (His)6-NSF was pretreated or not with 1 mM DEA-NONOate and then incubated with α-SNAP, VAMP-3, SNAP-23, and GST-syntaxin-4. ATP or ATP-γS 5 mM was added, the mixture was precipitated with glutathione-sepharose, and precipitated proteins were immunoblotted with antibodies as indicated. (D) NO inhibits NSF disassembly activity: dose response. Recombinant NSF was pre-treated with DEA-NONOate for 10 min, mixed with α-SNAP, and added to endothelial cell GST-SNARE fusion polypeptides as above. ATP 5 mM was added, GST-SNAREs were precipitated with glutathione-sepharose, and the precipitant was analyzed for the coprecipitation of NSF. (E) TAT-NSF peptides inhibit NSF ATPase activity. A colorimetric assay was used to measure the ATPase activity of recombinant NSF that had been treated with TAT-NSF peptides. (F) TAT-NSF peptides inhibit NSF disassembly activity. Recombinant NSF was pretreated with TAT-NSF peptides for 20 min and mixed with α-SNAP and endothelial cell GST-SNARE fusion polypeptides as above. ATP was added, GST-SNAREs were precipitated with glutathione-sepharose, and the precipitant was analyzed for the coprecipitation of NSF. (G) Exogenous NSF restores vWF exocytosis in endothelial cells treated with NO. HAEC were pretreated with 1 mM DEA-NONOate as above, permeabilized with SLO, incubated with recombinant NSF or nitrosylated recombinant NSF, stimulated with thrombin, and the amount of vWF in the media was measured (n = 3 ± SD, *p < 0.01 for NO versus NO + NSF).
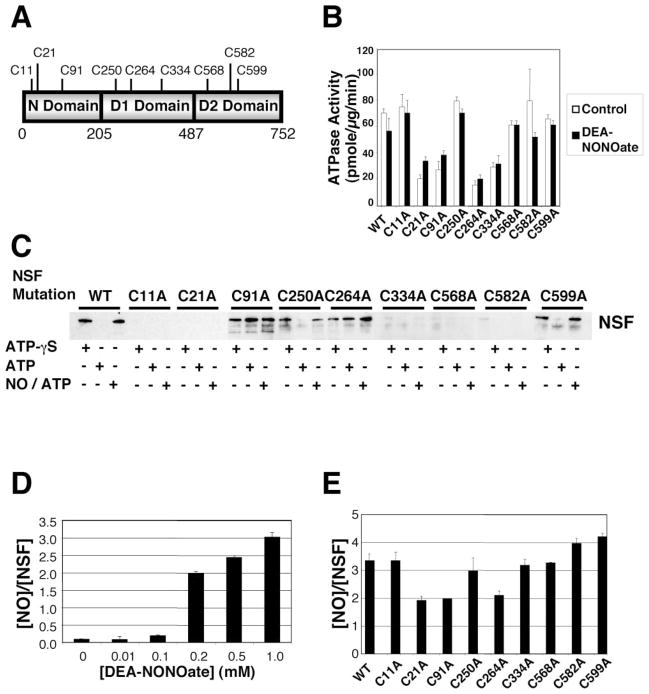
(A) Schematic of NSF. (B) Cysteine residues mediating ATPase activity of NSF. Wild-type NSF and mutant NSF were expressed and purified, treated with 1 mM DEA-NONOate or DEA, and the ATPase activity was measured as above. Mutagenesis of Cys at 21, 91, 264, and 334 partially inhibits NSF ATPase activity. NO does not inhibit ATPase activity of wild-type or mutant NSF (n = 3 ± SD). (C) Cysteine residues mediating disassembly activity of NSF. Wild-type NSF and mutant NSF (with mutated Cys residues) were expressed and purified, and disassembly activity was measured by the pull-down assay as above. Mutagenesis of Cys at 11, 21, 334, 568, and 582 inhibits NSF interaction with SNARE complex. Mutagenesis of Cys at 91 and 264 permits NSF interaction with SNARE complex but blocks NSF disassembly of SNARE complex. Mutagenesis of Cys at 250 and 599 has no effect on NSF interaction or disassembly. (D) Nitrosylation of NSF in vitro. DEA-NONOate was added to recombinant wild-type NSF, and the molar ratio of nitrosothiols per NSF molecule was measured by the Saville reaction. Each molecule of NSF can contain a maximum of three nitrosothiols (n = 3 ± SD). (E) Identification of nitrosylated cysteine residues in NSF. DEA-NONOate was added to recombinant NSF mutants, and the molar ratio of nitrosothiols per NSF molecule was measured by the Saville reaction. Mutation of C21 or C91 or C264 decreases the nitrosothiol content of nitrosylated NSF (n = 3–5 ± SD).

(A) NO inhibition of exocytosis is temporary. HAEC were pretreated with the NO donor SNAP 100 μM for 4 hr, washed to remove the NO donor, and then, at various times after NO treatment, thrombin was added for 1 hr, and the amount of vWF released into the media was measured (n = 3 ± SD). (B) NO nitrosylation of NSF is reversible. HAEC were pretreated with the NO donor SNAP 100 μM for 4 hr, washed to remove the NO donor, and cells were harvested at various times after treatment. (Top) Cell lysates were immunoprecipitated with antibody to nitrosocysteine and immunoblotted with antibody to NSF. (Bottom) Total cell lysates were immunoblotted with antibody to NSF (this experiment was repeated twice with similar results).
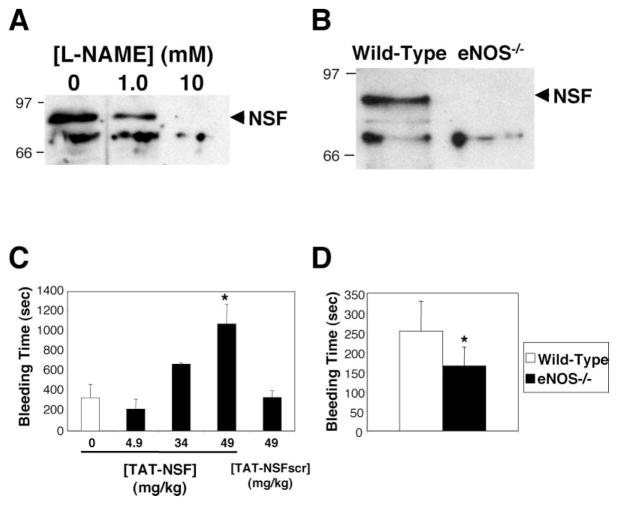
(A) Nitrosylation of NSF in human endothelial cells. HAEC were treated with media alone or the NOS inhibitor L-NAME. Nitrosothiols were biotinylated, precipitated with avidin-agarose, and precipitants were immunoblotted with antibody to NSF. L-NAME inhibits endogenous nitrosylation of NSF in human endothelial cells. (B) Nitrosylation of NSF in mice. Spleens from wild-type mice and mice lacking eNOS were harvested. Nitrosothiols were biotinylated, precipitated with avidin-agarose, and precipitants were immunoblotted with antibody to NSF. NSF is nitrosylated only in mice expressing eNOS. (C) Inhibition of NSF in mice prolongs bleeding time. Anesthetized mice were injected with PBS or the TAT-NSF222 peptide that inhibits NSF, and after 45 min, the distal tip of the tail was amputated and the bleeding time was measured. Bleeding that continued longer than 20 min was recorded as 20 min (n = 3–6 ± SD, *p < 0.01 for TAT-NSF versus TAT-NSFscr at 49 mg/kg). (D) Endogenous NO prolongs bleeding time in mice. The distal tip of the tail was amputated from wild-type and eNOS null mice, and the bleeding time was measured (n = 7–8 ± SD, *p = 0.01 for wild-type versus eNOS null).

(A) Exogenous NO inhibits histamine induced exocytosis in vitro. HAEC were pretreated with the NO donor SNAP 100 μM for 4 hr and then stimulated with histamine for 1 hr, and the amount of released vWF was measured as above (n = 2 ± SD, *p < 0.05 versus 0 μM). (B) Inhibition of endogenous NOS increases histamine induced platelet adherence to venules in mice. Mice were pretreated or not with 5 mg/kg L-NAME for 30 min and then transfused with calcein-AM labeled platelets. Intravital microscopy was used to visualize fluorescent platelets adhering to mesenteric venules 5 min after superfusion with 10 μl of 1 mM histamine. The dotted line indicates the edge of the mesenteric venule. The arrow points to a fluorescent platelet adhering to the vessel wall (bar = 50 μm). (C) Inhibition of endogenous NOS increases platelet adherence to venules in mice over time. Mice were pretreated (black circles) or not (white circles) with L-NAME and then transfused with calcein-AM labeled platelets as above. Intravital microscopy was used to visualize fluorescent platelets transiently adhering to mesenteric venules 0–20 min after superfusion with 10 μl of 1 mM histamine. Quantitative analysis of platelet adhesion as a function of time after the secretagogue application is shown (n = 4–5 mice ± SEM, *p < 0.05 and **p < 0.01 versus nontreated).
Comment in
-
S-nitrosylation of NSF controls membrane trafficking.Cell. 2003 Oct 17;115(2):127-9. doi: 10.1016/s0092-8674(03)00811-0. Cell. 2003. PMID: 14567907
References
-
- Andre P, Denis CV, Ware J, Saffaripour S, Hynes RO, Ruggeri ZM, Wagner DD. Platelets adhere to and translocate on von Willebrand factor presented by endothelium in stimulated veins. Blood. 2000;96:3322–3328. - PubMed
-
- Becker-Hapak M, McAllister SS, Dowdy SF. TAT-mediated protein transduction into mammalian cells. Methods. 2001;24:247–256. - PubMed
-
- Bonfanti R, Furie BC, Furie B, Wagner DD. PADGEM (GMP140) is a component of Weibel-Palade bodies of human endothelial cells. Blood. 1989;73:1109–1112. - PubMed
-
- Burgoyne RD, Morgan A. Analysis of regulated exocytosis in adrenal chromaffin cells: insights into NSF/SNAP/SNARE function. Bioessays. 1998;20:328–335. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- P01 HL065608/HL/NHLBI NIH HHS/United States
- R01 HL078635/HL/NHLBI NIH HHS/United States
- P01 HL056091/HL/NHLBI NIH HHS/United States
- R37 HL041002/HL/NHLBI NIH HHS/United States
- P01 HL65608/HL/NHLBI NIH HHS/United States
- RR07002/RR/NCRR NIH HHS/United States
- T32 RR007002/RR/NCRR NIH HHS/United States
- HL074945/HL/NHLBI NIH HHS/United States
- R01 HL063706/HL/NHLBI NIH HHS/United States
- K08 HL074945/HL/NHLBI NIH HHS/United States
- R37 HL41002/HL/NHLBI NIH HHS/United States
- P01 HL56091/HL/NHLBI NIH HHS/United States
- R01 HL63706-04/HL/NHLBI NIH HHS/United States
- R01 HL074061/HL/NHLBI NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
