

A critical role for OX40 in T cell-mediated immunopathology during lung viral infection
- PMID: 14568982
- PMCID: PMC2194232
- DOI: 10.1084/jem.20030351
A critical role for OX40 in T cell-mediated immunopathology during lung viral infection
Abstract
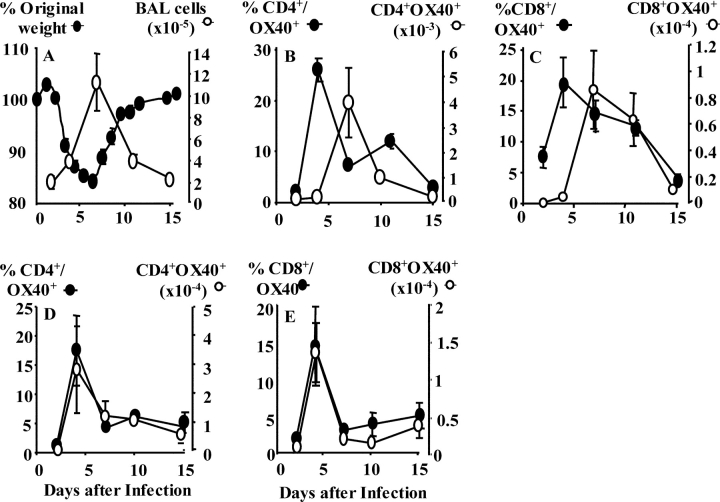
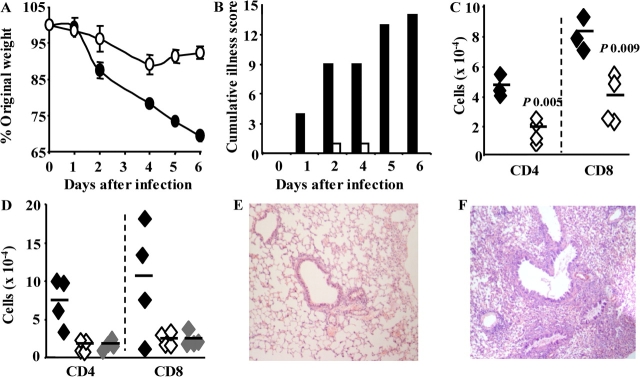
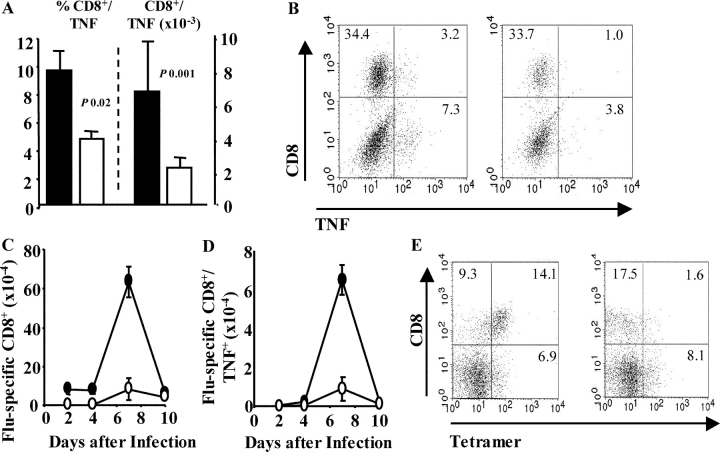
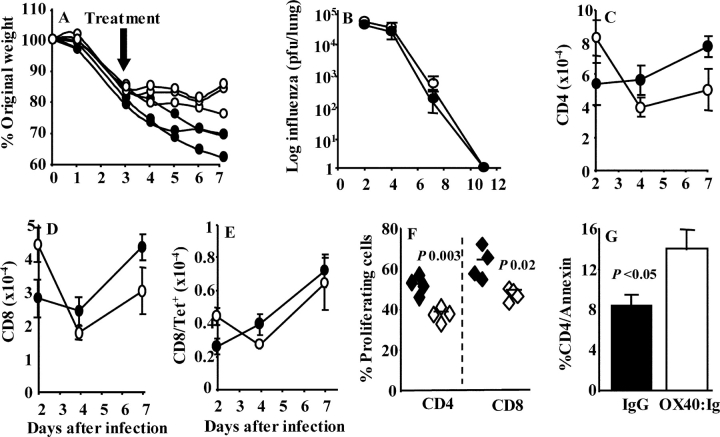
Respiratory infections are the third leading cause of death worldwide. Illness is caused by pathogen replication and disruption of airway homeostasis by excessive expansion of cell numbers. One strategy to prevent lung immune-mediated damage involves reducing the cellular burden. To date, antiinflammatory strategies have affected both antigen-specific and naive immune repertoires. Here we report a novel form of immune intervention that specifically targets recently activated T cells alone. OX40 (CD134) is absent on naive T cells but up-regulated 1-2 d after antigen activation. OX40-immunoglobulin fusion proteins block the interaction of OX40 with its ligand on antigen-presenting cells and eliminate weight loss and cachexia without preventing virus clearance. Reduced proliferation and enhanced apoptosis of lung cells accompanied the improved clinical phenotype. Manipulation of this late costimulatory pathway has clear therapeutic potential for the treatment of dysregulated lung immune responses.
Figures
References
-
- Murray, C.J., and A.D. Lopez. 1997. Global mortality, disability, and the contribution of risk factors: global burden of disease study. Lancet. 349:1436–1442. - PubMed
-
- Curwen, M., K. Dunnell, and J. Ashley. 1990. Hidden influenza deaths. Br. Med. J. 300:896.
-
- Doherty, P.C., D.J. Topham, R.A. Tripp, R.D. Cardin, J.W. Brooks, and P.G. Stevenson. 1997. Effector CD4+ and CD8+ T-cell mechanisms in the control of respiratory virus infections. Immunol. Rev. 159:105–117. - PubMed
-
- Hussell, T., A. Pennycook, and P.J.M. Openshaw. 2001. Inhibition of tumour necrosis factor reduces the severity of virus-specific lung immunopathology. Eur. J. Immunol. 31:2566–2573. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
