

Active sites of thioredoxin reductases: why selenoproteins?
- PMID: 14569031
- PMCID: PMC240667
- DOI: 10.1073/pnas.2134510100
Active sites of thioredoxin reductases: why selenoproteins?
Abstract
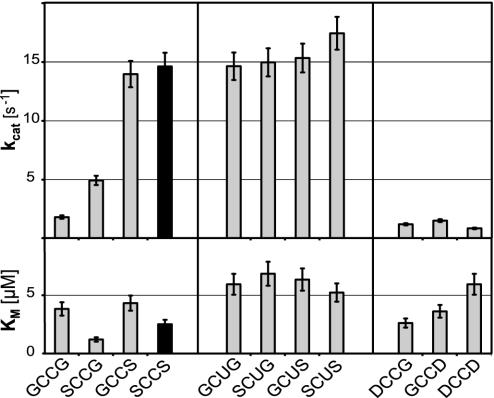
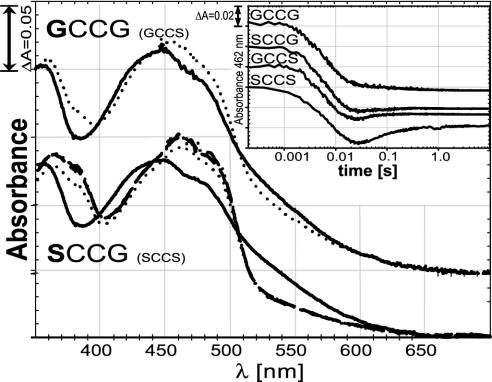
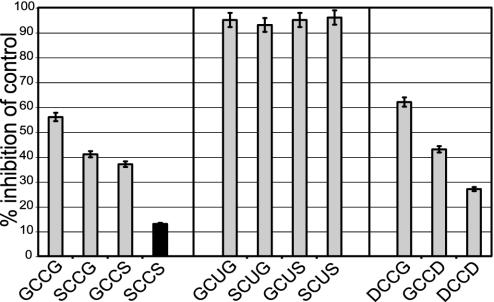
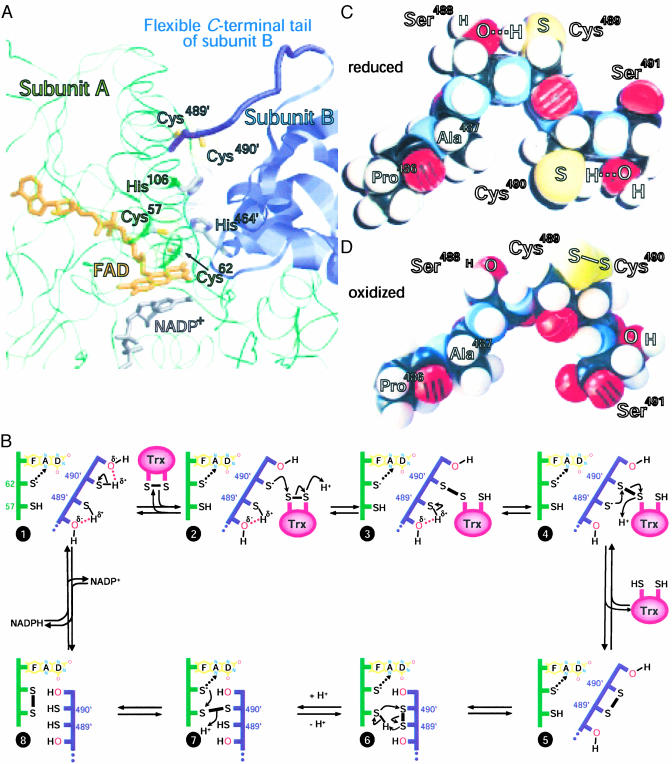
Selenium, an essential trace element for mammals, is incorporated into a selected class of selenoproteins as selenocysteine. All known isoenzymes of mammalian thioredoxin (Trx) reductases (TrxRs) employ selenium in the C-terminal redox center -Gly-Cys-Sec-Gly-COOH for reduction of Trx and other substrates, whereas the corresponding sequence in Drosophila melanogaster TrxR is -Ser-Cys-Cys-Ser-COOH. Surprisingly, the catalytic competence of these orthologous enzymes is similar, whereas direct Sec-to-Cys substitution of mammalian TrxR, or other selenoenzymes, yields almost inactive enzyme. TrxRs are therefore ideal for studying the biology of selenocysteine by comparative enzymology. Here we show that the serine residues flanking the C-terminal Cys residues of Drosophila TrxRs are responsible for activating the cysteines to match the catalytic efficiency of a selenocysteine-cysteine pair as in mammalian TrxR, obviating the need for selenium. This finding suggests that the occurrence of selenoenzymes, which implies that the organism is selenium-dependent, is not necessarily associated with improved enzyme efficiency. Our data suggest that the selective advantage of selenoenzymes is a broader range of substrates and a broader range of microenvironmental conditions in which enzyme activity is possible.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
