

Infection by Trypanosoma cruzi metacyclic forms deficient in gp82 but expressing a related surface molecule, gp30
- PMID: 14573635
- PMCID: PMC219548
- DOI: 10.1128/IAI.71.11.6184-6191.2003
Infection by Trypanosoma cruzi metacyclic forms deficient in gp82 but expressing a related surface molecule, gp30
Abstract
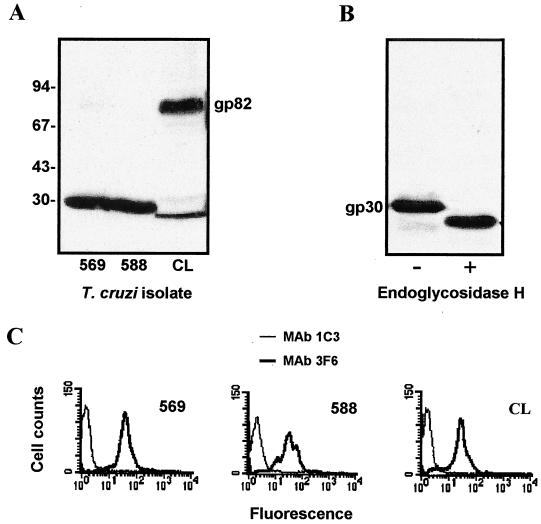
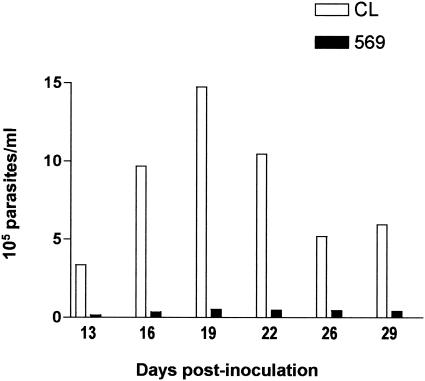
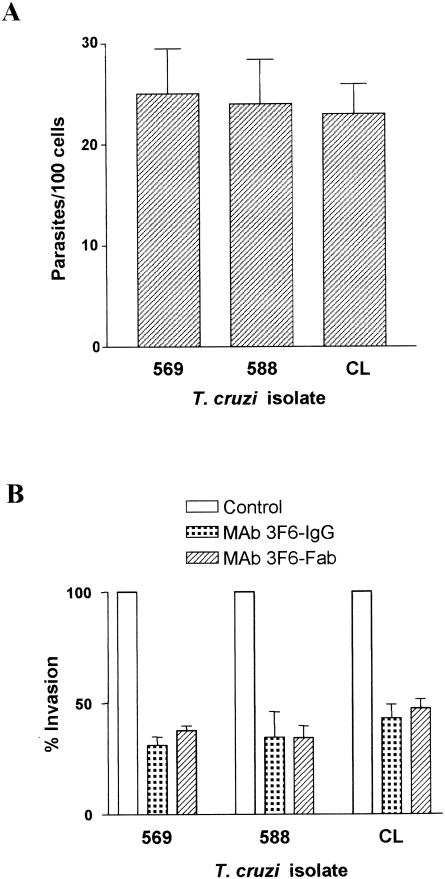
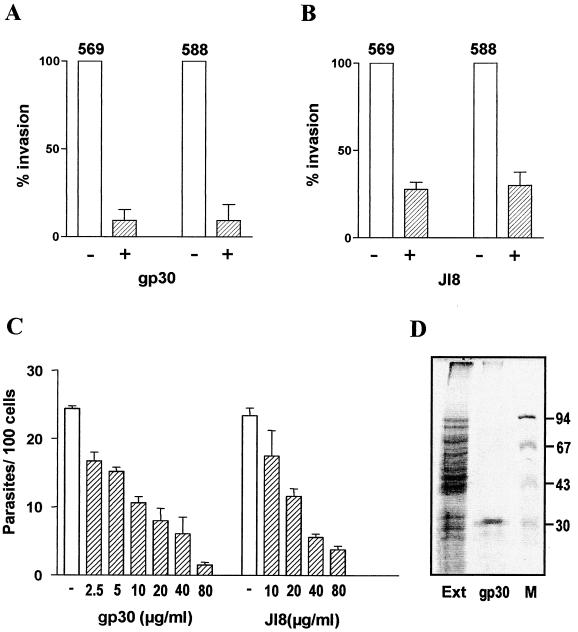
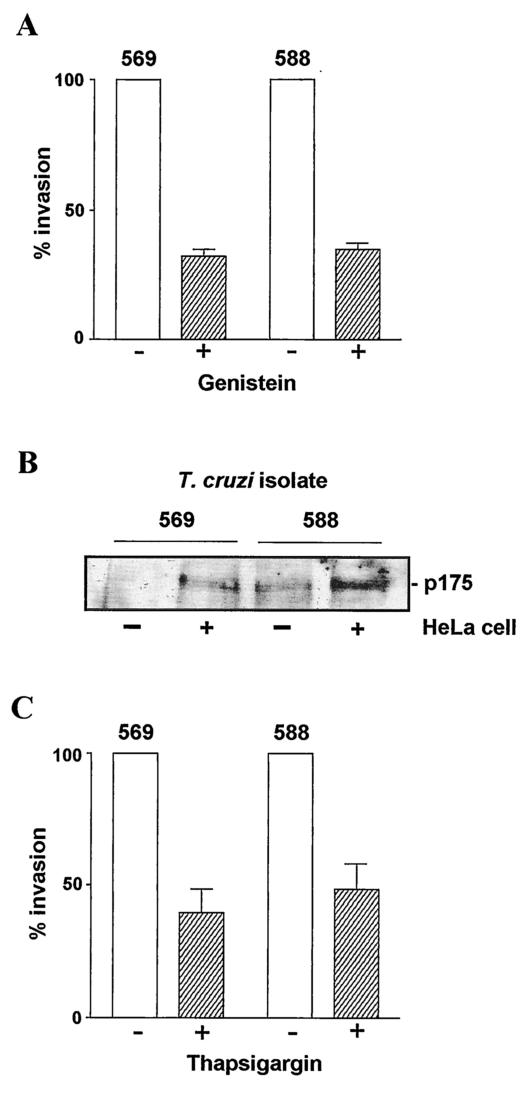
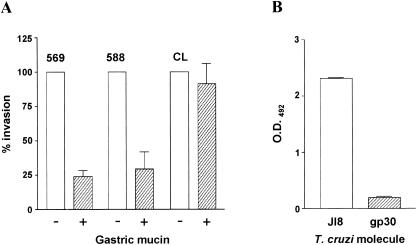
Trypanosoma cruzi metacyclic trypomastigotes invade and replicate in the gastric mucosal epithelium after oral infection. In this study we analyzed the process of infection by T. cruzi isolates deficient in the expression of gp82, the metacyclic stage-specific surface glycoprotein implicated in target cell entry in vitro and in promoting mucosal infection in mice after oral challenge. Mice infected by the oral route with metacyclic forms of gp82-deficient isolate 569 or 588 developed patent parasitemia but at greatly reduced levels compared to those infected with the gp82-expressing isolate CL. Metacyclic forms of both isolates expressed gp30, a surface glycoprotein detectable by monoclonal antibody (MAb) 3F6 directed to gp82. Otherwise, the gp82-deficient isolates displayed a surface profile similar to that of the CL isolate and also entered epithelial HeLa cells in a manner inhibitable by MAb 3F6 and dependent on the parasite signal transduction that involved the activation of protein tyrosine kinase and Ca(2+) mobilization from thapsigargin-sensitive stores. Like gp82, gp30 triggered the host cell Ca(2+) response required for parasite internalization. Purified gp30 and the recombinant gp82 inhibited HeLa cell invasion of metacyclic forms of isolates 569 and 588 by approximately 90 and approximately 70%, respectively. A cell invasion assay performed in the presence of gastric mucin, mimicking the in vivo infection, showed an inhibition of 70 to 75% in the internalization of gp82-deficient isolates but not of the CL isolate. The recombinant gp82 exhibited an adhesive capacity toward gastric mucin much higher than that of gp30. Taken together, our findings indicate that target cell entry of metacyclic trypomastigotes can be mediated either by gp82 or gp30 but that efficient mucosal infection depends on the expression of gp82.
Figures
References
-
- Akiyama, T., J. Ishida, S. Nakagawa, H. Ogawara, S. Watanabe, N. Itoh, M. Shibuya, and Y. Fukami. 1987. Genistein, a specific inhibitor of tyrosine-specific protein kinases. J. Biol. Chem. 262:5592-5595. - PubMed
-
- Brener, Z., and E. Chiari. 1963. Variações morfológicas observadas em diferentes amostras de Trypanosoma cruzi. Rev. Inst. Med. Trop. São Paulo 5:220-224. - PubMed
-
- Briones, M. R. S., R. P. Souto, B. S. Stolf, and B. Zingalez. 1999. The evolution of two Trypanosoma cruzi subgroups inferred from rRNA genes can be correlated with the interchange of American mammalian faunas in the Cenozoic and has implications to pathogenicity and host specificity. Mol. Biochem. Parasitol. 104:219-232. - PubMed
-
- Coura, J. R., A. C. V. Junqueira, O. Fernandes, S. A. S. Valente, and M. A. Miles. 2002. Emerging Chagas disease in Amazonian Brazil. Trends Parasitol. 18:171-176. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
