

Characterisation of new substrate specificities of Escherichia coli and Saccharomyces cerevisiae AP endonucleases
- PMID: 14576322
- PMCID: PMC275454
- DOI: 10.1093/nar/gkg812
Characterisation of new substrate specificities of Escherichia coli and Saccharomyces cerevisiae AP endonucleases
Abstract
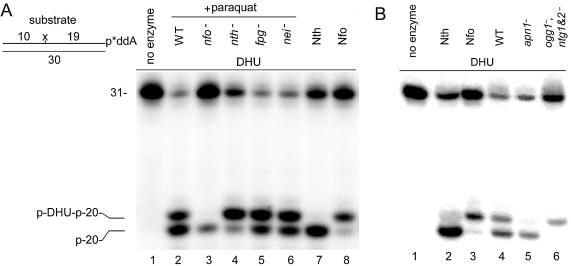
Despite the progress in understanding the base excision repair (BER) pathway it is still unclear why known mutants deficient in DNA glycosylases that remove oxidised bases are not sensitive to oxidising agents. One of the back-up repair pathways for oxidative DNA damage is the nucleotide incision repair (NIR) pathway initiated by two homologous AP endonucleases: the Nfo protein from Escherichia coli and Apn1 protein from Saccharomyces cerevisiae. These endonucleases nick oxidatively damaged DNA in a DNA glycosylase-independent manner, providing the correct ends for DNA synthesis coupled to repair of the remaining 5'-dangling nucleotide. NIR provides an advantage compared to DNA glycosylase-mediated BER, because AP sites, very toxic DNA glycosylase products, do not form. Here, for the first time, we have characterised the substrate specificity of the Apn1 protein towards 5,6-dihydropyrimidine, 5-hydroxy-2'-deoxyuridine and 2,6-diamino-4-hydroxy-5-N-methylformamidopyrimidine deoxynucleotide. Detailed kinetic comparisons of Nfo, Apn1 and various DNA glycosylases using different DNA substrates were made. The apparent K(m) and kcat/K(m) values of the reactions suggest that in vitro DNA glycosylase/AP lyase is somewhat more efficient than the AP endonuclease. However, in vivo, using cell-free extracts from paraquat-induced E.coli and from S.cerevisiae, we show that NIR is one of the major pathways for repair of oxidative DNA base damage.
Figures
Similar articles
-
Major oxidative products of cytosine are substrates for the nucleotide incision repair pathway.DNA Repair (Amst). 2007 Jan 4;6(1):8-18. doi: 10.1016/j.dnarep.2006.08.001. Epub 2006 Sep 15. DNA Repair (Amst). 2007. PMID: 16978929
-
Alpha-anomeric deoxynucleotides, anoxic products of ionizing radiation, are substrates for the endonuclease IV-type AP endonucleases.Biochemistry. 2004 Dec 7;43(48):15210-6. doi: 10.1021/bi049214+. Biochemistry. 2004. PMID: 15568813
-
Apurinic/apyrimidinic endonuclease Apn1 from Saccharomyces cerevisiae is recruited to the nucleotide incision repair pathway: Kinetic and structural features.Biochimie. 2018 Sep;152:53-62. doi: 10.1016/j.biochi.2018.06.012. Epub 2018 Jun 26. Biochimie. 2018. PMID: 29959063
-
Abasic sites in DNA: repair and biological consequences in Saccharomyces cerevisiae.DNA Repair (Amst). 2004 Jan 5;3(1):1-12. doi: 10.1016/j.dnarep.2003.10.002. DNA Repair (Amst). 2004. PMID: 14697754 Review.
-
Yeast base excision repair: interconnections and networks.Prog Nucleic Acid Res Mol Biol. 2001;68:29-39. doi: 10.1016/s0079-6603(01)68087-5. Prog Nucleic Acid Res Mol Biol. 2001. PMID: 11554305 Review.
Cited by
-
Selective Incision of the alpha-N-Methyl-Formamidopyrimidine Anomer by Escherichia coli Endonuclease IV.J Nucleic Acids. 2010 Jul 25;2010:850234. doi: 10.4061/2010/850234. J Nucleic Acids. 2010. PMID: 20798848 Free PMC article.
-
Yeast apurinic/apyrimidinic endonuclease Apn1 protects mammalian neuronal cell line from oxidative stress.J Neurochem. 2007 Jul;102(1):13-24. doi: 10.1111/j.1471-4159.2007.04490.x. Epub 2007 May 15. J Neurochem. 2007. PMID: 17506861 Free PMC article.
-
Recognition of a Clickable Abasic Site Analog by DNA Polymerases and DNA Repair Enzymes.Int J Mol Sci. 2022 Nov 1;23(21):13353. doi: 10.3390/ijms232113353. Int J Mol Sci. 2022. PMID: 36362137 Free PMC article.
-
Structural and Functional Characterization of a Unique AP Endonuclease From Deinococcus radiodurans.Front Microbiol. 2020 Jun 5;11:1178. doi: 10.3389/fmicb.2020.01178. eCollection 2020. Front Microbiol. 2020. PMID: 33117296 Free PMC article.
-
Mutational analysis of the damage-recognition and catalytic mechanism of human SMUG1 DNA glycosylase.Nucleic Acids Res. 2004 Oct 5;32(17):5291-302. doi: 10.1093/nar/gkh859. Print 2004. Nucleic Acids Res. 2004. PMID: 15466595 Free PMC article.
References
-
- Seeberg E., Eide,L. and Bjoras,M. (1995) The base excision repair pathway. Trends Biochem. Sci., 20, 391–397. - PubMed
-
- Lindahl T. (2001) Keynote: past, present and future aspects of base excision repair. Prog. Nucleic Acid Res. Mol. Biol., 68, xvii–xxx. - PubMed
-
- Scharer O.D. and Jiricny,J. (2001) Recent progress in the biology, chemistry and structural biology of DNA glycosylases. Bioessays, 23, 270–281. - PubMed
-
- Lindahl T. (1976) New class of enzymes acting on damaged DNA. Nature, 259, 64–66. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
