

Pathway shifts and thermal softening in temperature-coupled forced unfolding of spectrin domains
- PMID: 14581229
- PMCID: PMC1303605
- DOI: 10.1016/S0006-3495(03)74747-X
Pathway shifts and thermal softening in temperature-coupled forced unfolding of spectrin domains
Abstract
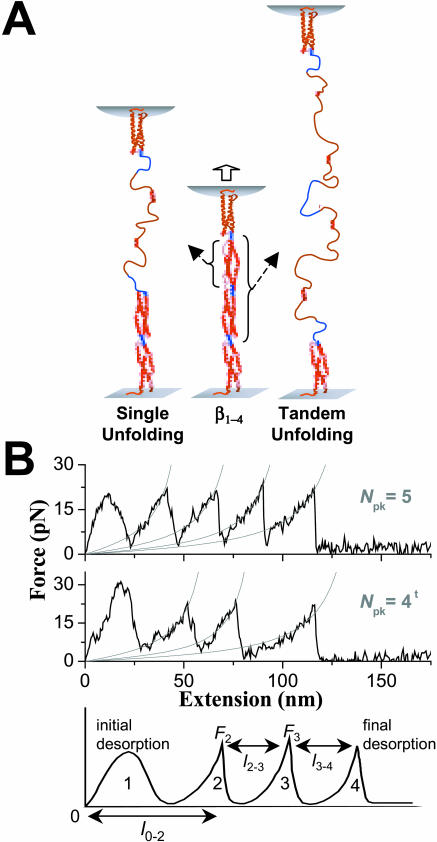
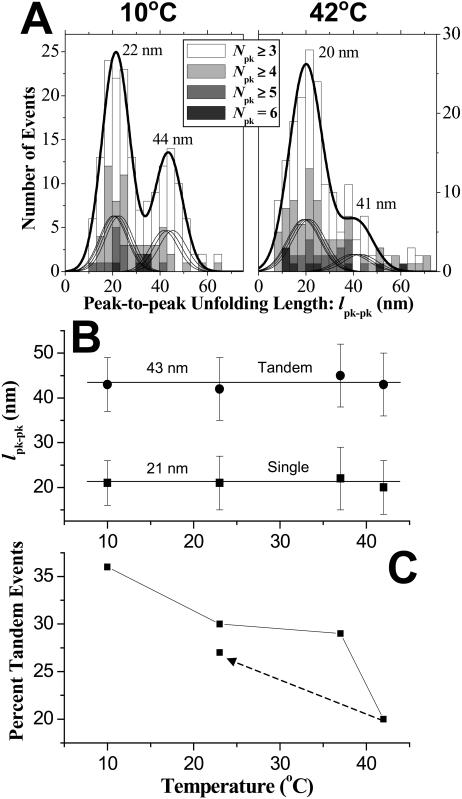
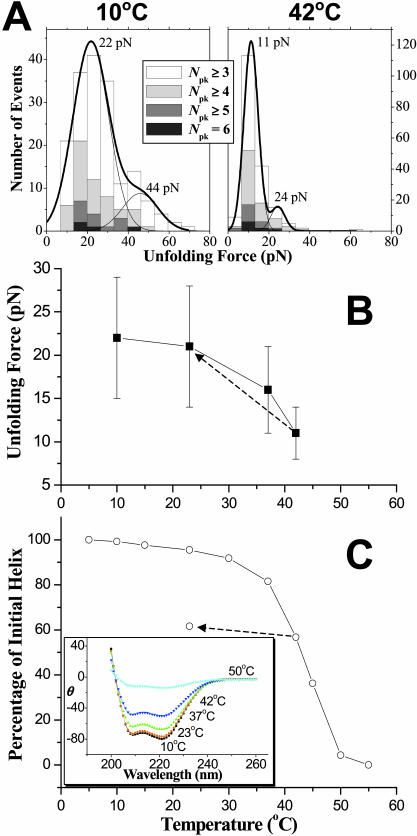
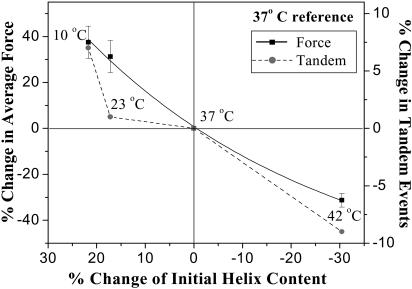
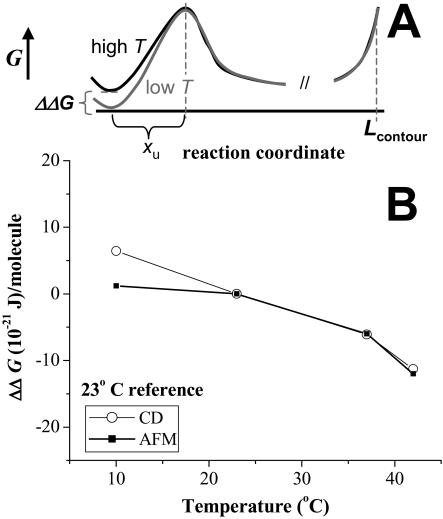
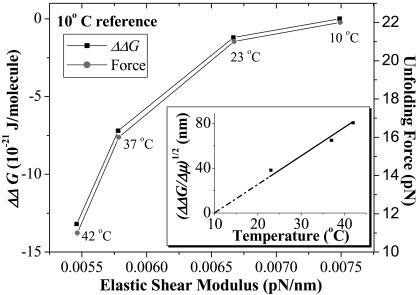
Pathways of unfolding a protein depend in principle on the perturbation-whether it is temperature, denaturant, or even forced extension. Widely-shared, helical-bundle spectrin repeats are known to melt at temperatures as low as 40-45 degrees C and are also known to unfold via multiple pathways as single molecules in atomic force microscopy. Given the varied roles of spectrin family proteins in cell deformability, we sought to determine the coupled effects of temperature on forced unfolding. Bimodal distributions of unfolding intervals are seen at all temperatures for the four-repeat beta(1-4) spectrin-an alpha-actinin homolog. The major unfolding length corresponds to unfolding of a single repeat, and a minor peak at twice the length corresponds to tandem repeats. Increasing temperature shows fewer tandem events but has no effect on unfolding intervals. As T approaches T(m), however, mean unfolding forces in atomic force microscopy also decrease; and circular dichroism studies demonstrate a nearly proportional decrease of helical content in solution. The results imply a thermal softening of a helical linker between repeats which otherwise propagates a helix-to-coil transition to adjacent repeats. In sum, structural changes with temperature correlate with both single-molecule unfolding forces and shifts in unfolding pathways.
Figures
References
-
- Altmann, S., R. Grunberg, P. Lenne, J. Ylanne, A. Raae, K. Herbert, M. Saraste, M. Nilges, and J. Horber. 2002. Pathways and intermediates in forced unfolding of spectrin repeats. Structure. 10:1085–1096. - PubMed
-
- Bell, G. I. 1978. Models for the specific adhesion of cells to cells. Science. 200:618–627. - PubMed
-
- Cantor, C. R., and P. R. Schimmel. 1980. Biophysical Chemistry, Part III—The Behavior of Biological Macromolecules. W.H. Freeman and Co., New York.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
