

A dual-functional paramyxovirus F protein regulatory switch segment: activation and membrane fusion
- PMID: 14581458
- PMCID: PMC2173521
- DOI: 10.1083/jcb.200305130
A dual-functional paramyxovirus F protein regulatory switch segment: activation and membrane fusion
Abstract
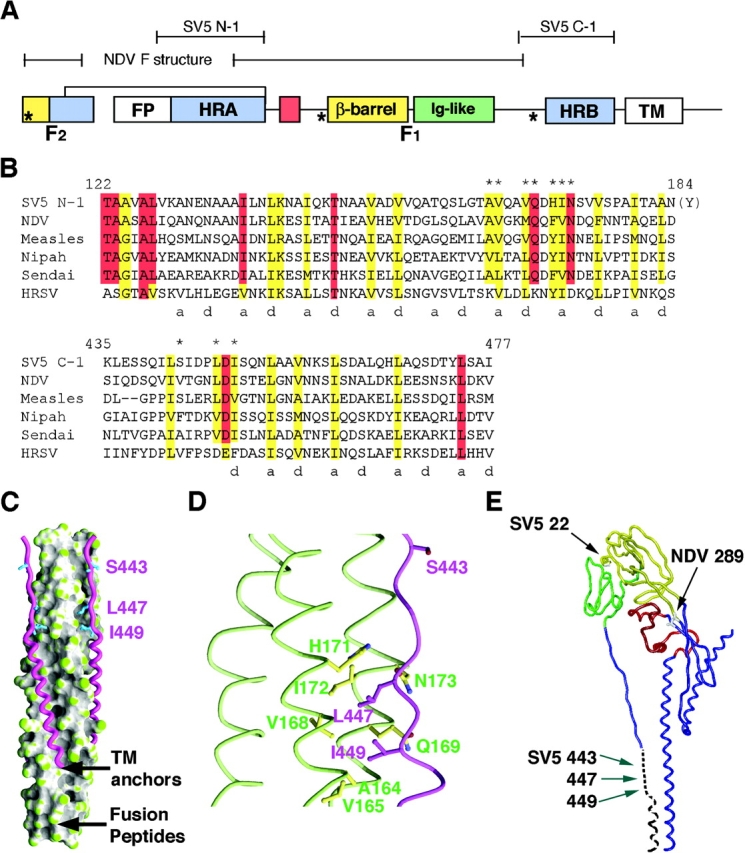
Many viral fusion-mediating glycoproteins couple alpha-helical bundle formation to membrane merger, but have different methods for fusion activation. To study paramyxovirus-mediated fusion, we mutated the SV5 fusion (F) protein at conserved residues L447 and I449, which are adjacent to heptad repeat (HR) B and bind to a prominent cavity in the HRA trimeric coiled coil in the fusogenic six-helix bundle (6HB) structure. These analyses on residues L447 and I449, both in intact F protein and in 6HB, suggest a metamorphic region around these residues with dual structural roles. Mutation of L447 and I449 to aliphatic residues destabilizes the 6HB structure and attenuates fusion activity. Mutation of L447 and I449 to aromatic residues also destabilizes the 6HB structure despite promoting hyperactive fusion, indicating that 6HB stability alone does not dictate fusogenicity. Thus, residues L447 and I449 adjacent to HRB in paramyxovirus F have distinct roles in fusion activation and 6HB formation, suggesting this region is involved in a conformational switch.
Figures






References
-
- Baker, K.A., R.E. Dutch, R.A. Lamb, and T.S. Jardetzky. 1999. Structural basis for paramyxovirus-mediated membrane fusion. Mol. Cell. 3:309–319. - PubMed
-
- Buckland, R., E. Malvoisin, P. Beauverger, and F. Wild. 1992. A leucine zipper structure present in the measles virus fusion protein is not required for its tetramerization but is essential for fusion. J. Gen. Virol. 73:1703–1707. - PubMed
-
- Bullough, P.A., F.M. Hughson, J.J. Skehel, and D.C. Wiley. 1994. Structure of influenza haemagglutinin at the pH of membrane fusion. Nature. 371:37–43. - PubMed
-
- Calder, L.J., L. Gonzalez-Reyes, B. Garcia-Barreno, S.A. Wharton, J.J. Skehel, D.C. Wiley, and J.A. Melero. 2000. Electron microscopy of the human respiratory syncytial virus fusion protein and complexes that it forms with monoclonal antibodies. Virology. 271:122–131. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
