

The crystal structures of severe acute respiratory syndrome virus main protease and its complex with an inhibitor
- PMID: 14585926
- PMCID: PMC263746
- DOI: 10.1073/pnas.1835675100
The crystal structures of severe acute respiratory syndrome virus main protease and its complex with an inhibitor
Abstract
A newly identified severe acute respiratory syndrome coronavirus (SARS-CoV), is the etiological agent responsible for the outbreak of SARS. The SARS-CoV main protease, which is a 33.8-kDa protease (also called the 3C-like protease), plays a pivotal role in mediating viral replication and transcription functions through extensive proteolytic processing of two replicase polyproteins, pp1a (486 kDa) and pp1ab (790 kDa). Here, we report the crystal structures of the SARS-CoV main protease at different pH values and in complex with a specific inhibitor. The protease structure has a fold that can be described as an augmented serine-protease, but with a Cys-His at the active site. This series of crystal structures, which is the first, to our knowledge, of any protein from the SARS virus, reveal substantial pH-dependent conformational changes, and an unexpected mode of inhibitor binding, providing a structural basis for rational drug design.
Figures
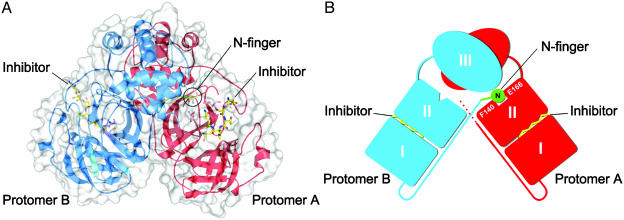
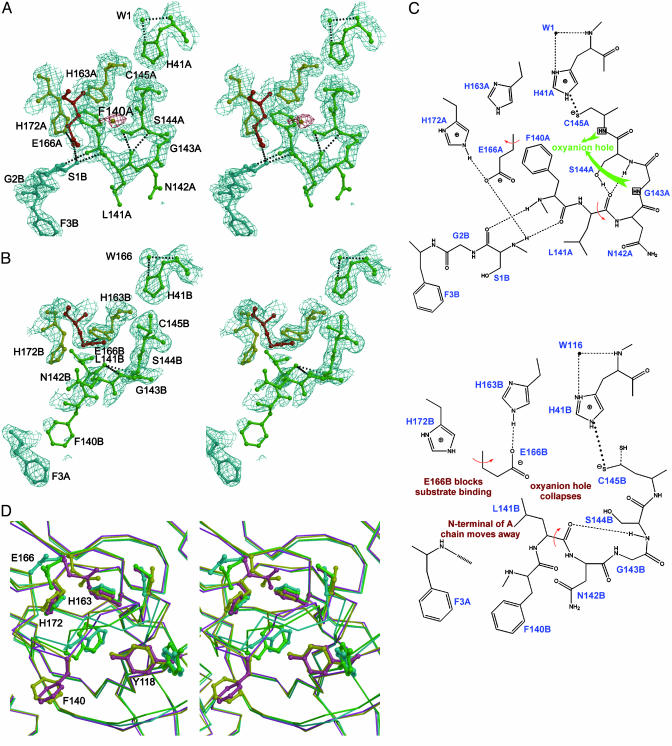
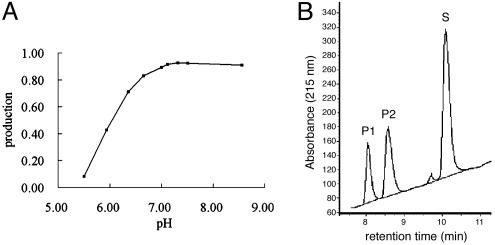
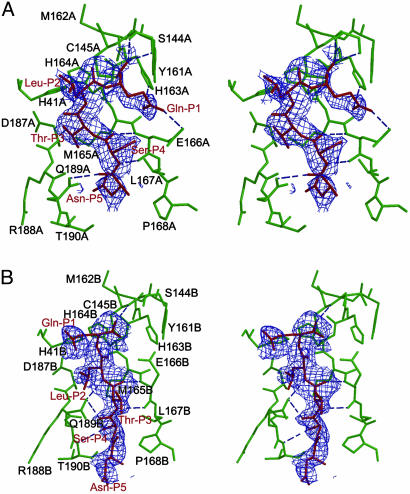
References
-
- Drosten, C., Gunther, S., Preiser, W., van der Werf, S., Brodt, H. R., Becker, S., Rabenau, H., Panning, M., Kolesnikova, L., Fouchier, R. A., et al. (2003) N. Engl. J. Med. 348, 1967-1976. - PubMed
-
- Ksiazek, T. G., Erdman, D., Goldsmith, C. S., Zaki, S. R., Peret, T., Emery, S., Tong, S., Urbani, C., Comer, J. A., Lim, W., et al. (2003) N. Engl. J. Med. 348, 1953-1966. - PubMed
-
- Marra, M. A., Jones, S. J., Astell, C. R., Holt, R. A., Brooks-Wilson, A., Butterfield, Y. S., Khattra, J., Asano, J. K., Barber, S. A., Chan, S. Y., et al. (2003) Science 300, 1399-1404. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
