

Discrete functions of TRAF1 and TRAF2 in Drosophila melanogaster mediated by c-Jun N-terminal kinase and NF-kappaB-dependent signaling pathways
- PMID: 14585960
- PMCID: PMC262421
- DOI: 10.1128/MCB.23.22.7982-7991.2003
Discrete functions of TRAF1 and TRAF2 in Drosophila melanogaster mediated by c-Jun N-terminal kinase and NF-kappaB-dependent signaling pathways
Abstract
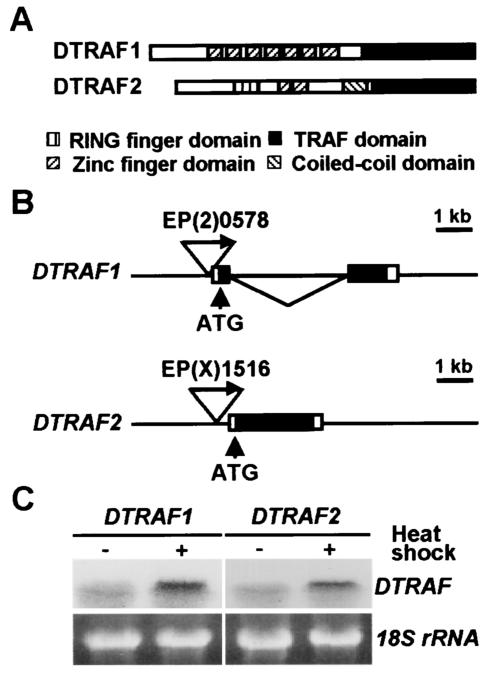
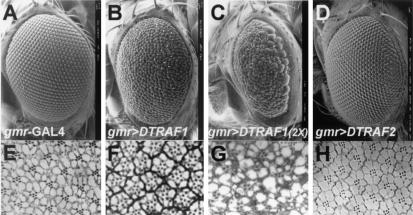
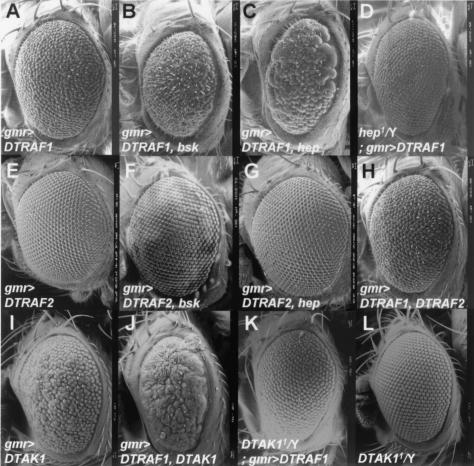
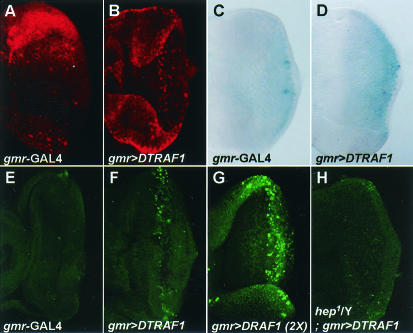
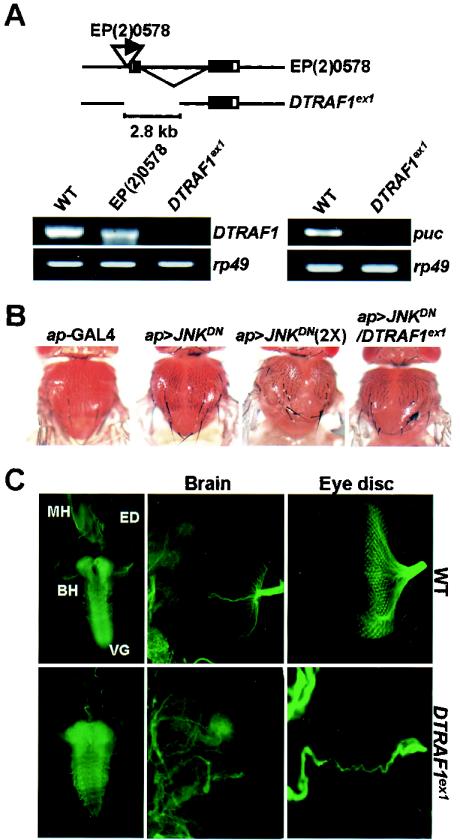
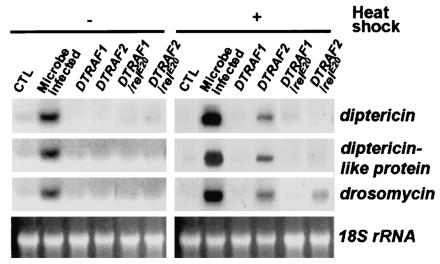
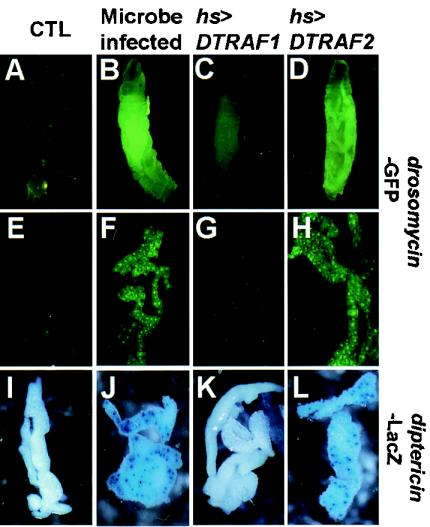
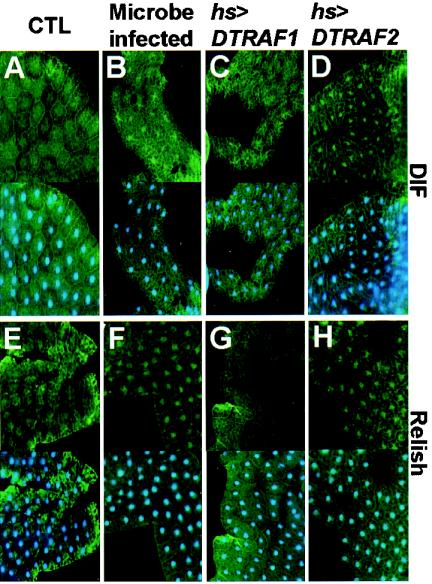
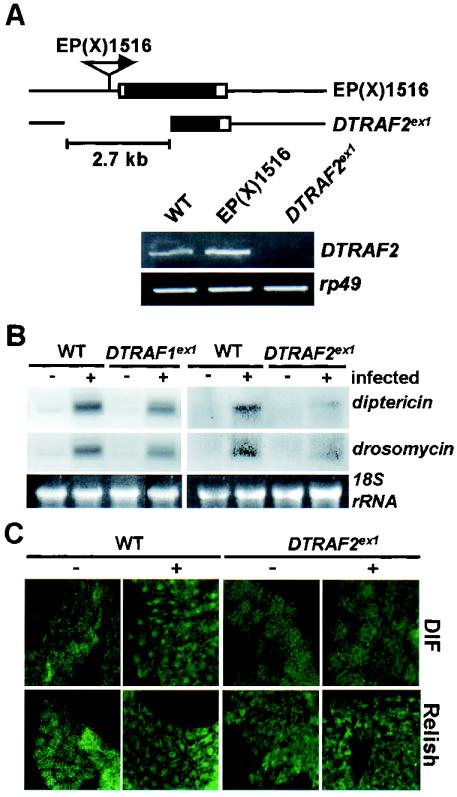
Two Drosophila tumor necrosis factor receptor-associated factors (TRAF), DTRAF1 and DTRAF2, are proposed to have similar functions with their mammalian counterparts as a signal mediator of cell surface receptors. However, their in vivo functions and related signaling pathways are not fully understood yet. Here, we show that DTRAF1 is an in vivo regulator of c-Jun N-terminal kinase (JNK) pathway in Drosophila melanogaster. Ectopic expression of DTRAF1 in the developing eye induced apoptosis, thereby causing a rough-eye phenotype. Further genetic interaction analyses revealed that the apoptosis in the eye imaginal disc and the abnormal eye morphogenesis induced by DTRAF1 are dependent on JNK and its upstream kinases, Hep and DTAK1. In support of these results, DTRAF1-null mutant showed a remarkable reduction in JNK activity with an impaired development of imaginal discs and a defective formation of photosensory neuron arrays. In contrast, DTRAF2 was demonstrated as an upstream activator of nuclear factor-kappaB (NF-kappaB). Ectopic expression of DTRAF2 induced nuclear translocation of two Drosophila NF-kappaBs, DIF and Relish, consequently activating the transcription of the antimicrobial peptide genes diptericin, diptericin-like protein, and drosomycin. Consistently, the null mutant of DTRAF2 showed immune deficiencies in which NF-kappaB nuclear translocation and antimicrobial gene transcription against microbial infection were severely impaired. Collectively, our findings demonstrate that DTRAF1 and DTRAF2 play pivotal roles in Drosophila development and innate immunity by differentially regulating the JNK- and the NF-kappaB-dependent signaling pathway, respectively.
Figures
References
-
- Adachi-Yamada, T., K. Fujimura-Kamada, Y. Nishida, and K. Matsumoto. 1999. Distortion of proheximodistal information causes JNK-dependent apoptosis in Drosophila wing. Nature 400:166-169. - PubMed
-
- Adachi-Yamada, T., M. Nakamura, K. Irie, Y. Tomoyasu, Y. Sano, E. Mori, S. Goto, N. Ueno, Y. Nishida, and K. Matsumoto. 1999. p38 mitogen-activated protein kinase can be involved in transforming growth factor beta superfamily signal transduction in Drosophila wing morphogenesis. Mol. Cell. Biol. 19:2322-2329. - PMC - PubMed
-
- Arch, R. H., R. W. Gedrich, and C. B. Thompson. 1998. Tumor necrosis factor receptor-associated factors (TRAFs): a family of adapter proteins that regulates life and death. Genes Dev. 12:2821-2830. - PubMed
-
- Bulet, P., J. L. Dimarcq, C. Hetru, M. Lagueux, M. Charlet, G. Hegy, A. Van Dorsselaer, and J. A. Hoffmann. 1993. A novel inducible antibacterial peptide of Drosophila carries an O-glycosylated substitution. J. Biol. Chem. 268:14893-14897. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous