

Phosphorylation-elicited quaternary changes of GA binding protein in transcriptional activation
- PMID: 14585962
- PMCID: PMC262348
- DOI: 10.1128/MCB.23.22.8008-8018.2003
Phosphorylation-elicited quaternary changes of GA binding protein in transcriptional activation
Abstract
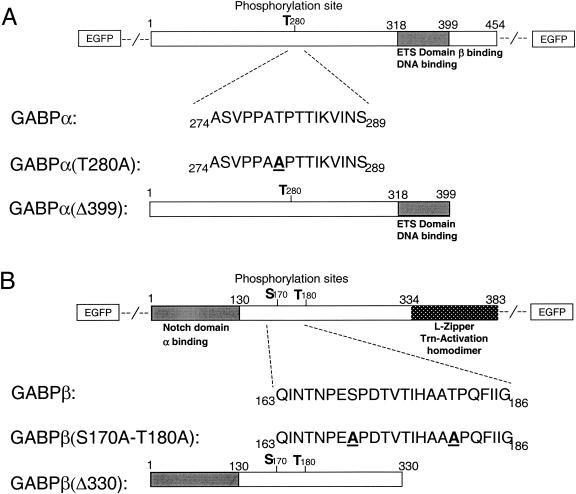
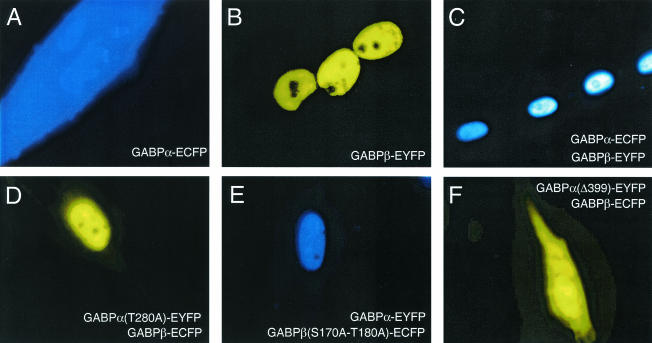
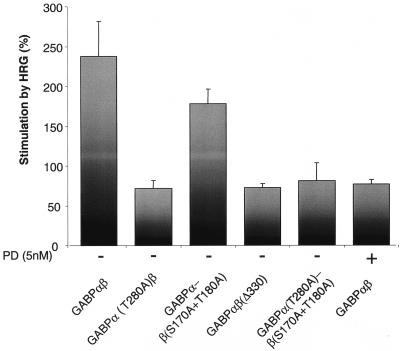
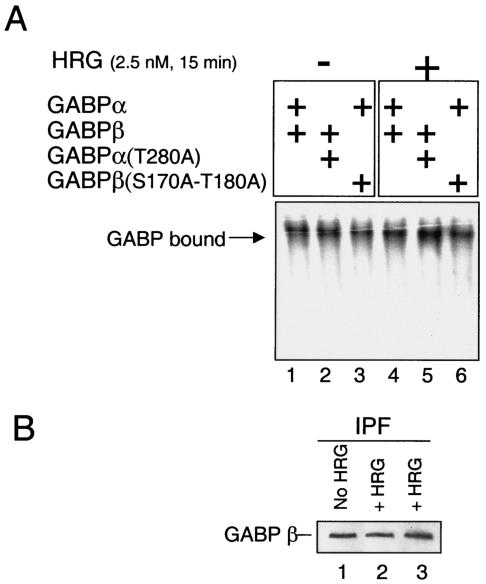
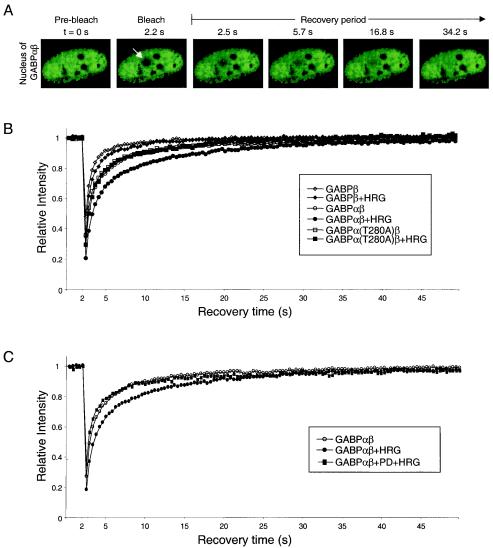
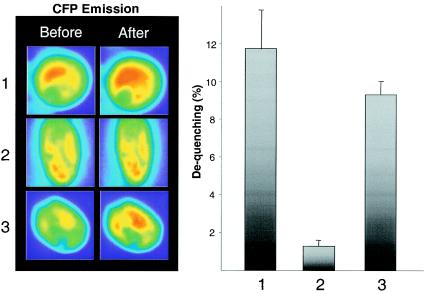
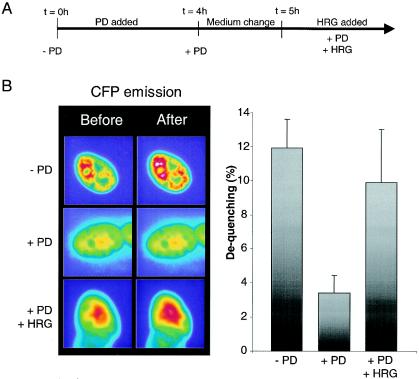
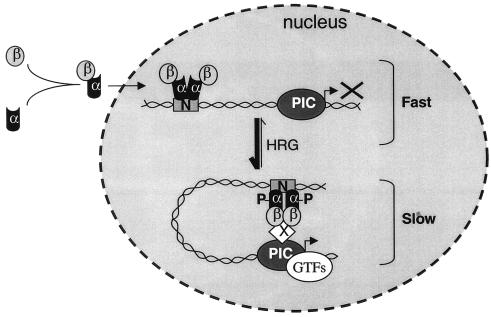
Enrichment of nicotinic acetylcholine receptors (nAChR) on the tip of the subjunctional folds of the postsynaptic membrane is a central event in the development of the vertebrate neuromuscular junction. This is attained, in part, through a selective transcription in the subsynaptic nuclei, and it has recently been shown that the GA binding protein (GABP) plays an important role in this compartmentalized expression. The neural factor heregulin (HRG) activates nAChR transcription in cultured cells by stimulating a signaling cascade of protein kinases. Hence, it is speculated that GABP becomes activated by phosphorylation, but the mechanism has remained elusive. To fully understand the consequences of GABP phosphorylation, we examined the effect of heregulin-elicited GABP phosphorylation on cellular localization, DNA binding, transcription, and mobility. We demonstrate that HRG-elicited phosphorylation dramatically changes the transcriptional activity and mobility of GABP. While phosphorylation of GABPbeta seems to be dispensable for these changes, phosphorylation of GABPalpha is crucial. Using fluorescence resonance energy transfer, we furthermore showed that phosphorylation of threonine 280 in GABPalpha triggers reorganizations of the quaternary structure of GABP. Taken together, these results support a model in which phosphorylation-elicited structural changes of GABP enable engagement in certain interactions leading to transcriptional activation.
Figures
Similar articles
-
Neuregulin-1-stimulated phosphorylation of GABP in skeletal muscle cells.Biochemistry. 2001 May 1;40(17):5306-12. doi: 10.1021/bi002649m. Biochemistry. 2001. PMID: 11318655
-
Neuregulin-1 induces acetylcholine receptor transcription in the absence of GABPalpha phosphorylation.J Neurosci Res. 2008 Apr;86(5):982-91. doi: 10.1002/jnr.21563. J Neurosci Res. 2008. PMID: 17975830
-
Implication of a multisubunit Ets-related transcription factor in synaptic expression of the nicotinic acetylcholine receptor.EMBO J. 1998 Jun 1;17(11):3078-90. doi: 10.1093/emboj/17.11.3078. EMBO J. 1998. PMID: 9606190 Free PMC article.
-
GA-binding protein transcription factor: a review of GABP as an integrator of intracellular signaling and protein-protein interactions.Blood Cells Mol Dis. 2004 Jan-Feb;32(1):143-54. doi: 10.1016/j.bcmd.2003.09.005. Blood Cells Mol Dis. 2004. PMID: 14757430 Review.
-
Targeting transcription to the neuromuscular synapse.Neuron. 2001 Jul 19;31(1):15-22. doi: 10.1016/s0896-6273(01)00353-1. Neuron. 2001. PMID: 11498047 Review.
Cited by
-
KDM5-mediated activation of genes required for mitochondrial biology is necessary for viability in Drosophila.Development. 2023 Nov 1;150(21):dev202024. doi: 10.1242/dev.202024. Epub 2023 Nov 6. Development. 2023. PMID: 37800333 Free PMC article.
-
Basic helix-loop-helix factors recruit nuclear factor I to enhance expression of the NaV 1.4 Na+ channel gene.Biochim Biophys Acta. 2007 Nov-Dec;1769(11-12):649-58. doi: 10.1016/j.bbaexp.2007.08.004. Epub 2007 Sep 14. Biochim Biophys Acta. 2007. PMID: 17936922 Free PMC article.
-
Formation of IL-7Ralphahigh and IL-7Ralphalow CD8 T cells during infection is regulated by the opposing functions of GABPalpha and Gfi-1.J Immunol. 2008 Apr 15;180(8):5309-19. doi: 10.4049/jimmunol.180.8.5309. J Immunol. 2008. PMID: 18390712 Free PMC article.
-
Identification of a crucial site for synoviolin expression.Mol Cell Biol. 2005 Aug;25(16):7344-56. doi: 10.1128/MCB.25.16.7344-7356.2005. Mol Cell Biol. 2005. PMID: 16055742 Free PMC article.
-
GA-binding protein is dispensable for neuromuscular synapse formation and synapse-specific gene expression.Mol Cell Biol. 2007 Jul;27(13):5040-6. doi: 10.1128/MCB.02228-06. Epub 2007 May 7. Mol Cell Biol. 2007. PMID: 17485447 Free PMC article.
References
-
- Batchelor, A. H., D. E. Piper, F. C. de la Brousse, S. L. McKnight, and C. Wolberger. 1998. The structure of GABPalpha/beta: an ETS domain-ankyrin repeat heterodimer bound to DNA. Science 279:1037-1041. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous