

The FACT complex travels with elongating RNA polymerase II and is important for the fidelity of transcriptional initiation in vivo
- PMID: 14585989
- PMCID: PMC262413
- DOI: 10.1128/MCB.23.22.8323-8333.2003
The FACT complex travels with elongating RNA polymerase II and is important for the fidelity of transcriptional initiation in vivo
Erratum in
- Mol Cell Biol. 2004 Jul;24(14):6536
Abstract
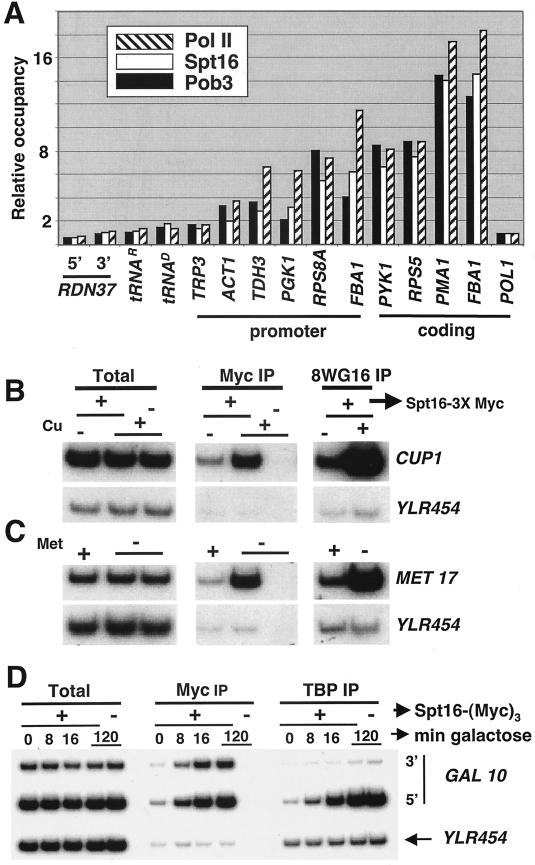
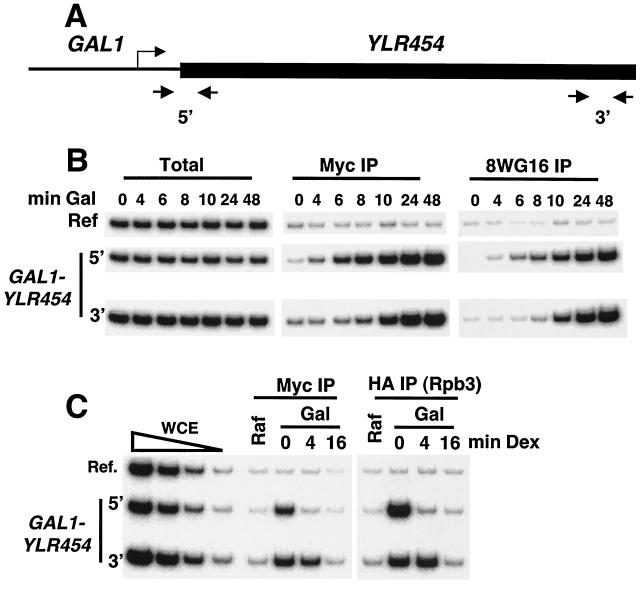
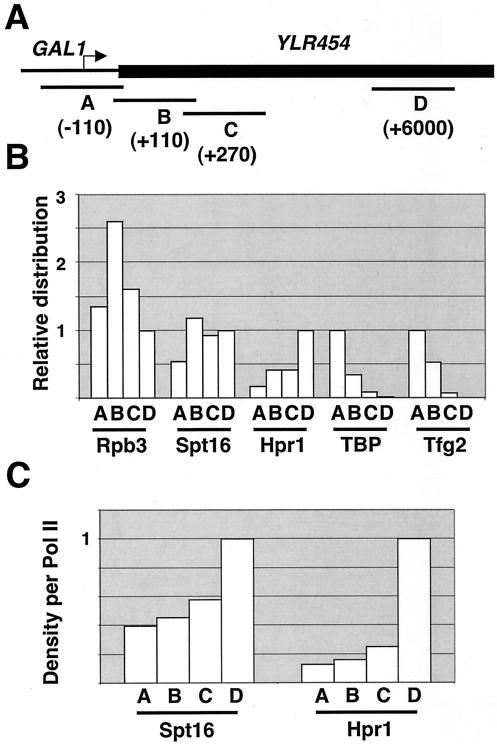
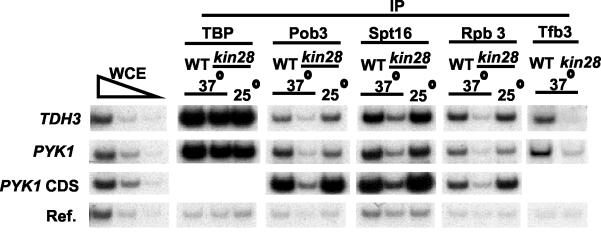
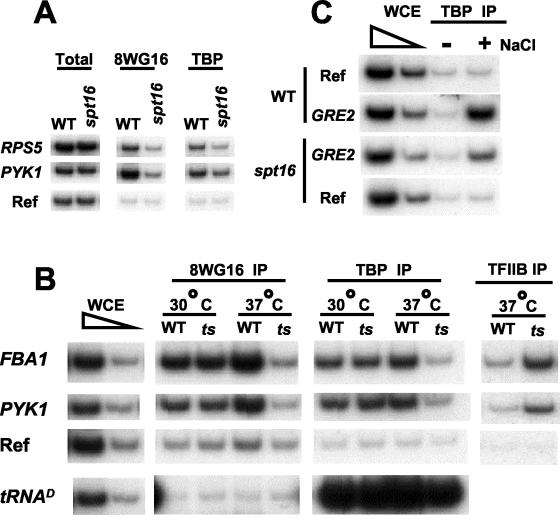
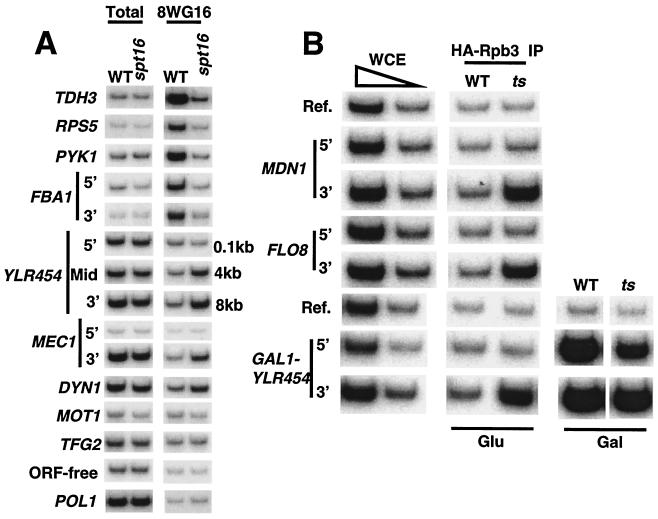
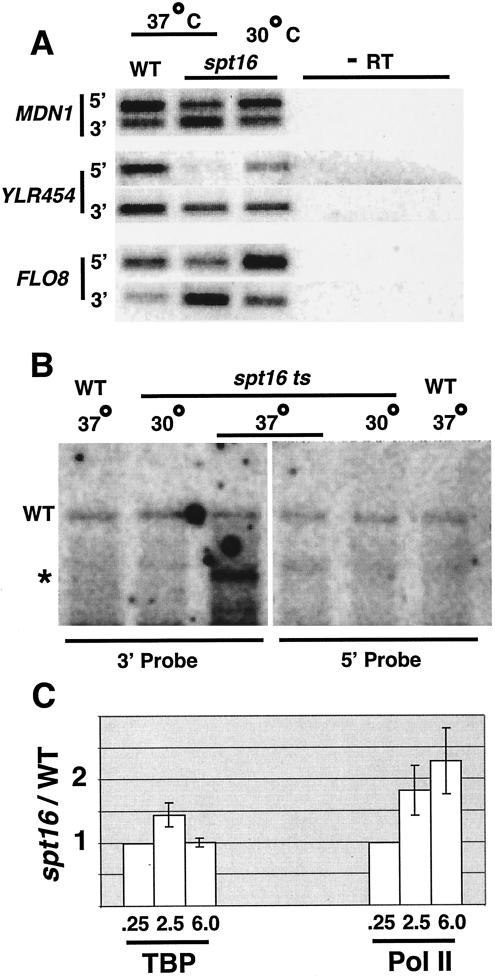
The FACT complex facilitates transcription on chromatin templates in vitro, and it has been functionally linked to nucleosomes and putative RNA polymerase II (Pol II) elongation factors. In Saccharomyces cerevisiae cells, FACT specifically associates with active Pol II genes in a TFIIH-dependent manner and travels across the gene with elongating Pol II. Conditional inactivation of the FACT subunit Spt16 results in increased Pol II density, transcription, and TATA-binding protein (TBP) occupancy in the 3' portion of certain coding regions, indicating that FACT suppresses inappropriate initiation from cryptic promoters within coding regions. Conversely, loss of Spt16 activity reduces the association of TBP, TFIIB, and Pol II with normal promoters. Thus, FACT is required for wild-type cells to restrict initiation to normal promoters, thereby ensuring that only appropriate mRNAs are synthesized. We suggest that FACT contributes to the fidelity of Pol II transcription by linking the processes of initiation and elongation.
Figures
References
-
- Bentley, D. L., and M. Groudine. 1986. A block to elongation is largely responsible for decreased transcription of c-myc in differentiated HL60 cells. Nature 321:702-706. - PubMed
-
- Brewster, N. K., G. C. Johnston, and R. A. Singer. 1998. Characterization of the CP complex, an abundant dimer of Cdc68 and Pob3 proteins that regulates yeast transcriptional activation and chromatin repression. J. Biol. Chem. 273:21972-21979. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials