

Extinction requires new RNA and protein synthesis and the soma of the cell right pedal dorsal 1 in Lymnaea stagnalis
- PMID: 14586013
- PMCID: PMC6740901
- DOI: 10.1523/JNEUROSCI.23-30-09842.2003
Extinction requires new RNA and protein synthesis and the soma of the cell right pedal dorsal 1 in Lymnaea stagnalis
Abstract
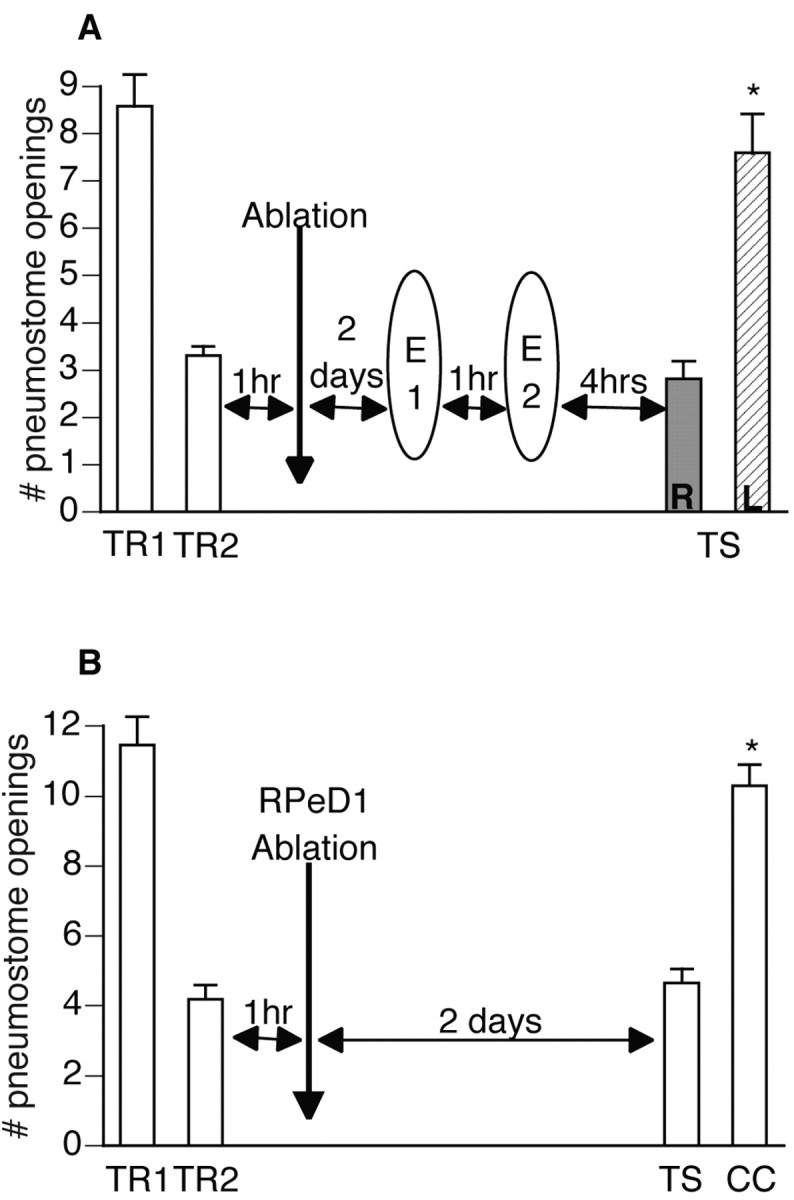
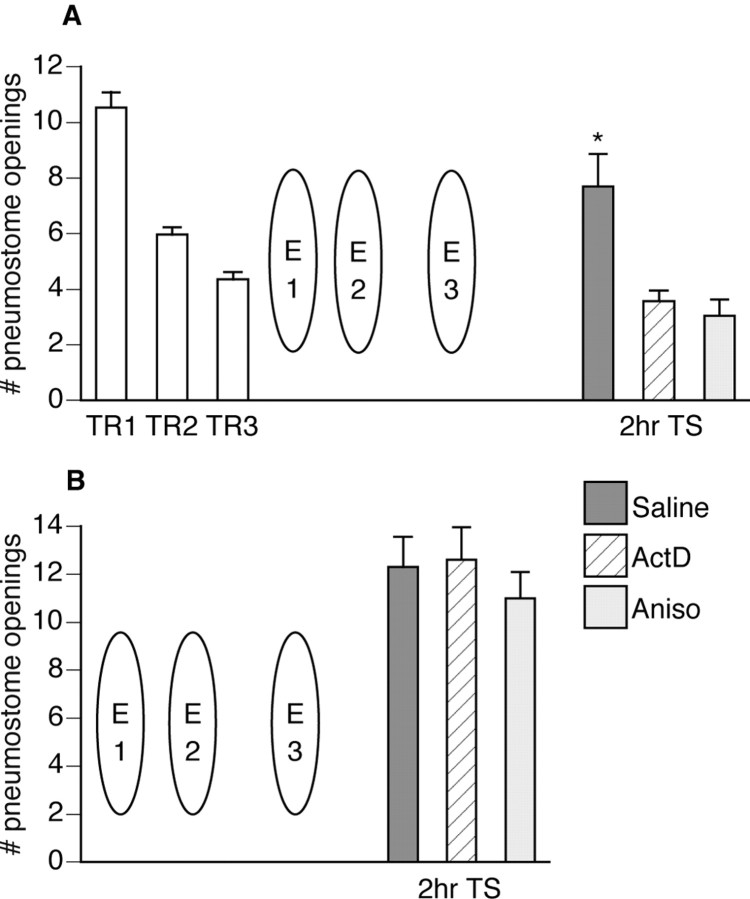
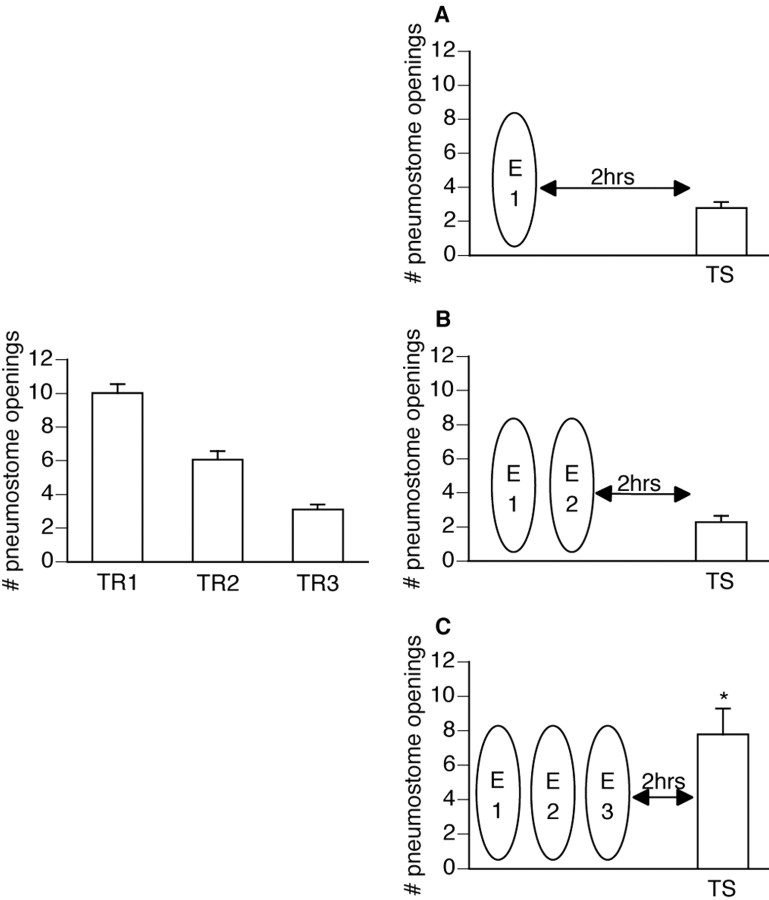
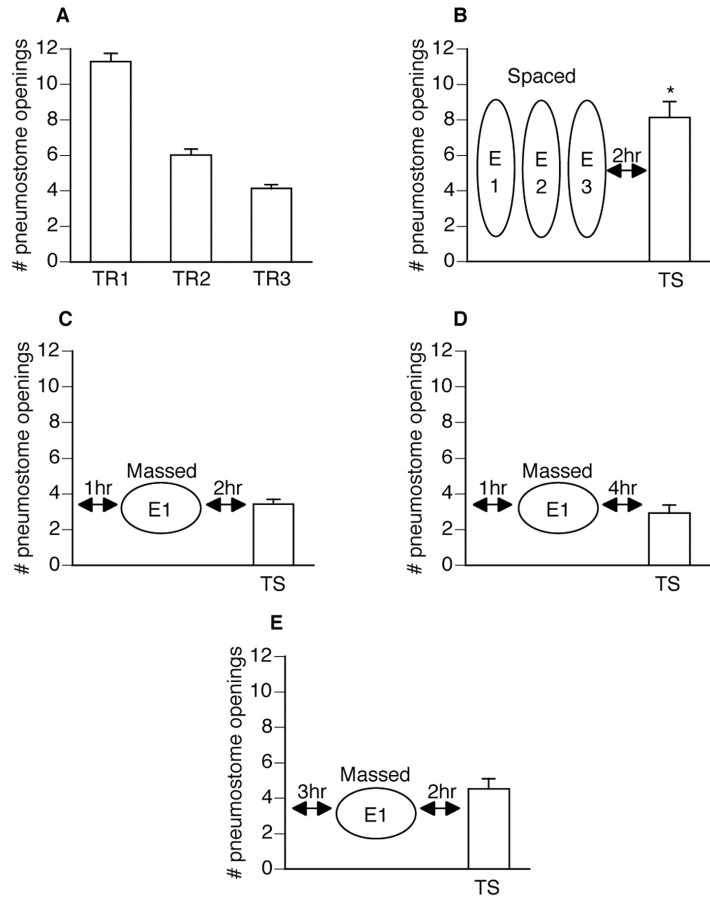
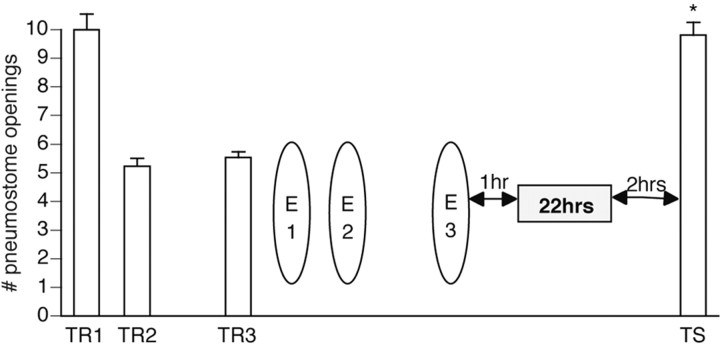
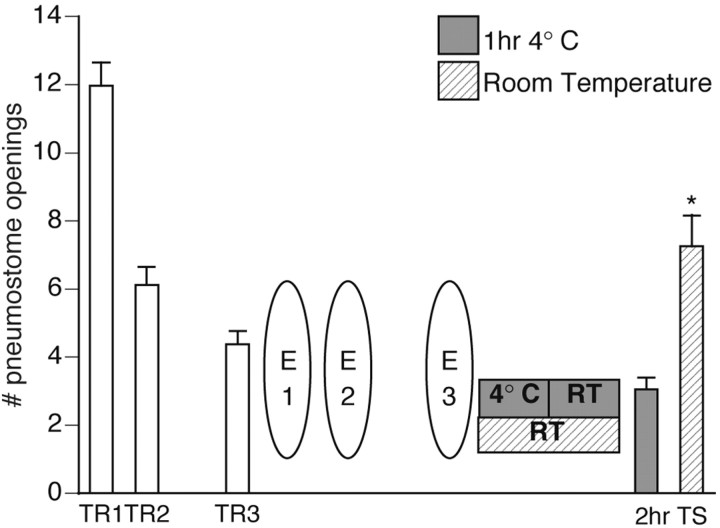
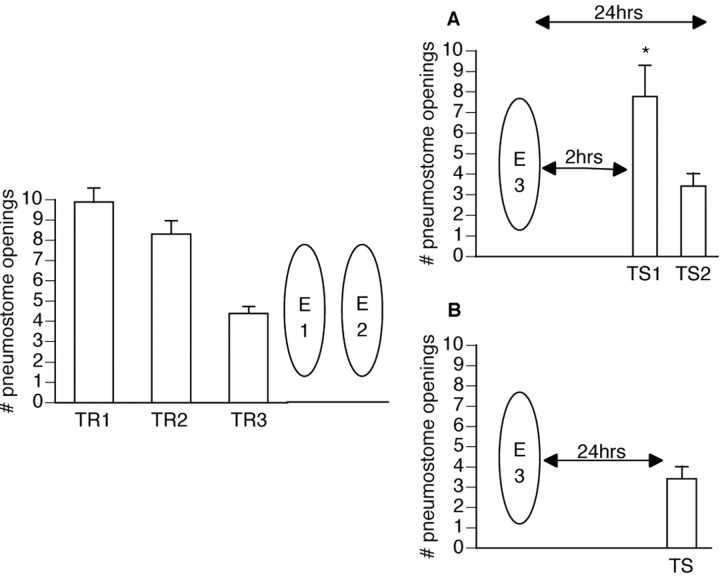
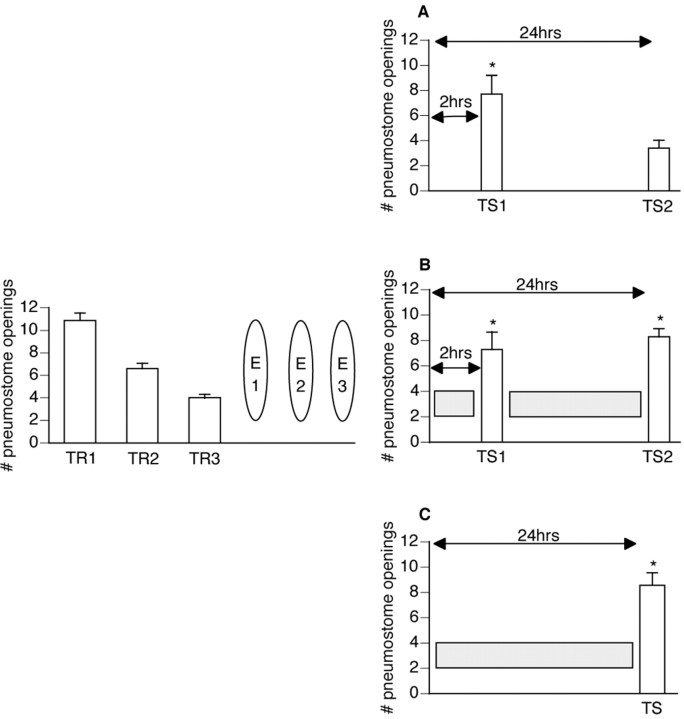
Lymnaea stagnalis were operantly conditioned to not perform aerial respiratory behavior. This learned response was subsequently extinguished. Here, we show that spaced extinction training is more effective than massed extinction training, in addition to the occurrence of spontaneous recovery. We also find evidence of a critical period within the first hour after extinction training in which new RNA and protein synthesis must occur for a memory of extinction training to be established. The memory for extinction training can also be extended using cooling and by preventing aerial respiration from occurring after extinction training. In addition, we demonstrate that memory formation of extinction training requires the soma of the cell right pedal dorsal 1, a cell that we have previously shown to be necessary for long-term memory consolidation and reconsolidation. This finding implies that the events that lead to the formation of extinction memory occur in the same cell that is responsible for long-term memory of operant conditioning. All of these data are consistent with the hypothesis that, during extinction, a new associative memory is being formed and that this new memory covers up, but does not abolish, the "old" memory.
Figures
Similar articles
-
Reconsolidation of a long-term memory in Lymnaea requires new protein and RNA synthesis and the soma of right pedal dorsal 1.J Neurosci. 2003 Sep 3;23(22):8034-40. doi: 10.1523/JNEUROSCI.23-22-08034.2003. J Neurosci. 2003. PMID: 12954865 Free PMC article.
-
Methamphetamine enhances memory of operantly conditioned respiratory behavior in the snail Lymnaea stagnalis.J Exp Biol. 2010 Jun 15;213(Pt 12):2055-65. doi: 10.1242/jeb.042820. J Exp Biol. 2010. PMID: 20511519
-
Intermediate and long-term memories of associative learning are differentially affected by transcription versus translation blockers in Lymnaea.J Exp Biol. 2003 May;206(Pt 10):1605-13. doi: 10.1242/jeb.00301. J Exp Biol. 2003. PMID: 12682092
-
Learning, memory and a respiratory central pattern generator.Comp Biochem Physiol A Mol Integr Physiol. 1999 Nov;124(3):265-74. doi: 10.1016/s1095-6433(99)00114-2. Comp Biochem Physiol A Mol Integr Physiol. 1999. PMID: 10665379 Review.
-
A molluscan model system in the search for the engram.J Physiol Paris. 2003 Jan;97(1):69-76. doi: 10.1016/j.jphysparis.2003.10.008. J Physiol Paris. 2003. PMID: 14706692 Review.
Cited by
-
Persistent disruption of an established morphine conditioned place preference.J Neurosci. 2006 Mar 15;26(11):3010-20. doi: 10.1523/JNEUROSCI.4818-05.2006. J Neurosci. 2006. PMID: 16540579 Free PMC article.
-
Function and mechanisms of memory destabilization and reconsolidation after retrieval.Proc Jpn Acad Ser B Phys Biol Sci. 2020;96(3):95-106. doi: 10.2183/pjab.96.008. Proc Jpn Acad Ser B Phys Biol Sci. 2020. PMID: 32161213 Free PMC article. Review.
-
Epicatechin, a component of dark chocolate, enhances memory formation if applied during the memory consolidation period.Commun Integr Biol. 2016 Jul 27;9(4):e1205772. doi: 10.1080/19420889.2016.1205772. eCollection 2016 Jul-Aug. Commun Integr Biol. 2016. PMID: 27574544 Free PMC article.
-
The participation of NMDA receptors, PKC, and MAPK in the formation of memory following operant conditioning in Lymnaea.Mol Brain. 2010 Aug 31;3:24. doi: 10.1186/1756-6606-3-24. Mol Brain. 2010. PMID: 20807415 Free PMC article.
-
Extinction: [corrected] does it or doesn't it? The requirement of altered gene activity and new protein synthesis.Biol Psychiatry. 2006 Aug 15;60(4):344-51. doi: 10.1016/j.biopsych.2006.05.038. Biol Psychiatry. 2006. PMID: 16919523 Free PMC article. Review.
References
-
- Berman DE, Dudai Y ( 2001) Memory extinction, learning anew, and learning the new: dissociations in the molecular machinery of learning in cortex. Science 291: 2417–2419. - PubMed
-
- Bouton ME ( 1993) Context, time and memory retrieval in the interference paradigms of Pavlovian conditioning. Psychol Bull 114: 80–99. - PubMed
-
- Cartford MC, Gohl EB, Singson M, Lavond DG ( 1997) The effects of reversible inactivation of the red nucleus on learning-related and auditory-evoked unit activity in the pontine nuclei of classically conditioned rabbits. Learn Mem 3: 519–531. - PubMed
-
- Davis HP, Squire LR ( 1984) Protein synthesis and memory: a review. Psychol Bull 96: 518–559. - PubMed
-
- Davis M, Myers KM ( 2002) The role of glutamate and gamma-aminobutyric acid in fear extinction: clinical implications for exposure therapy. Biol Psychiatry 52: 998–1007. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources