

Exportin 6: a novel nuclear export receptor that is specific for profilin.actin complexes
- PMID: 14592989
- PMCID: PMC275422
- DOI: 10.1093/emboj/cdg565
Exportin 6: a novel nuclear export receptor that is specific for profilin.actin complexes
Abstract
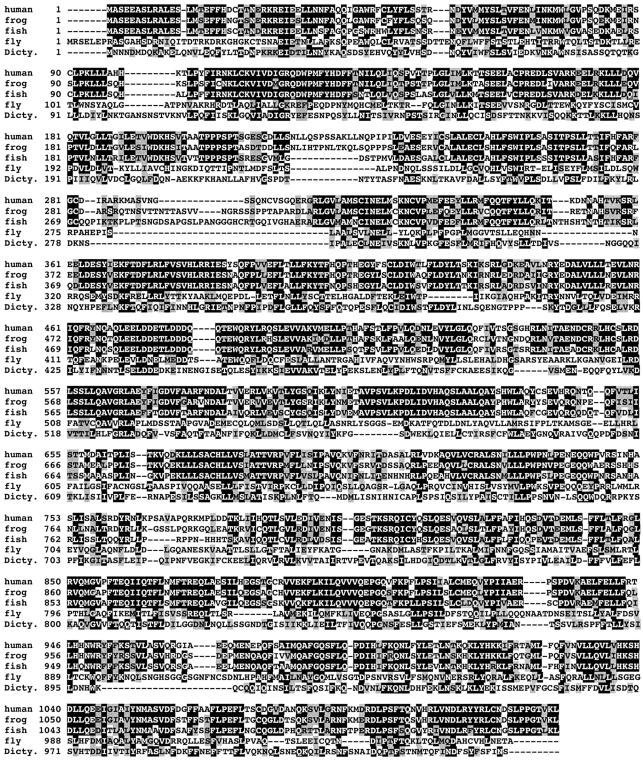
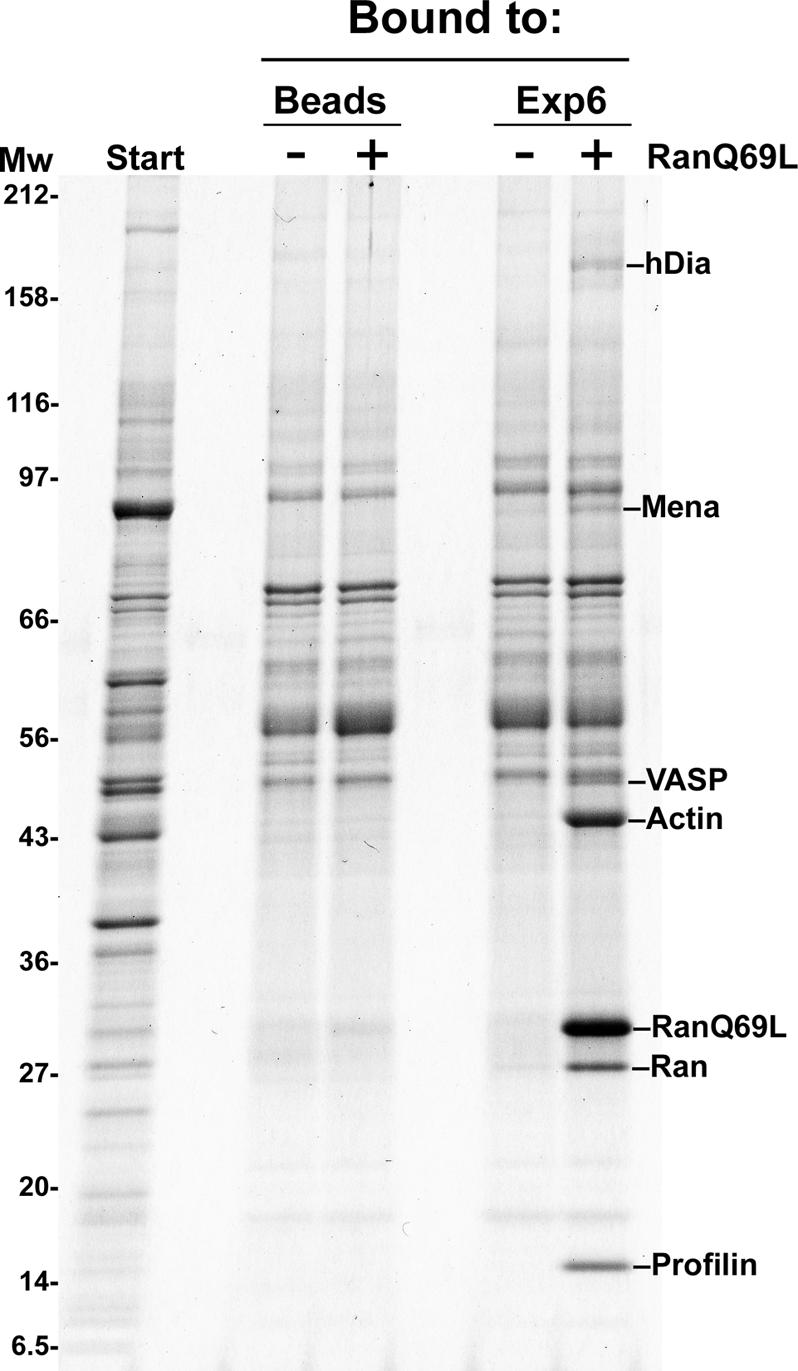
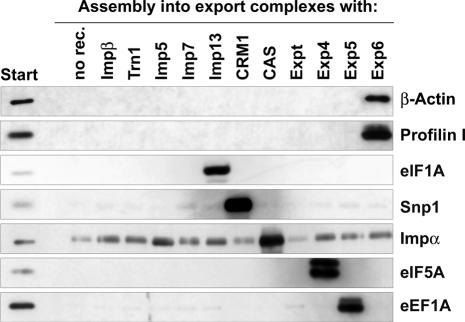
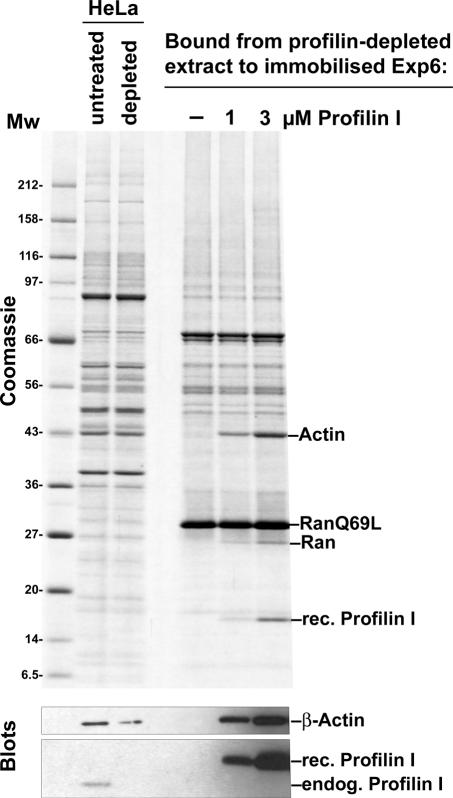
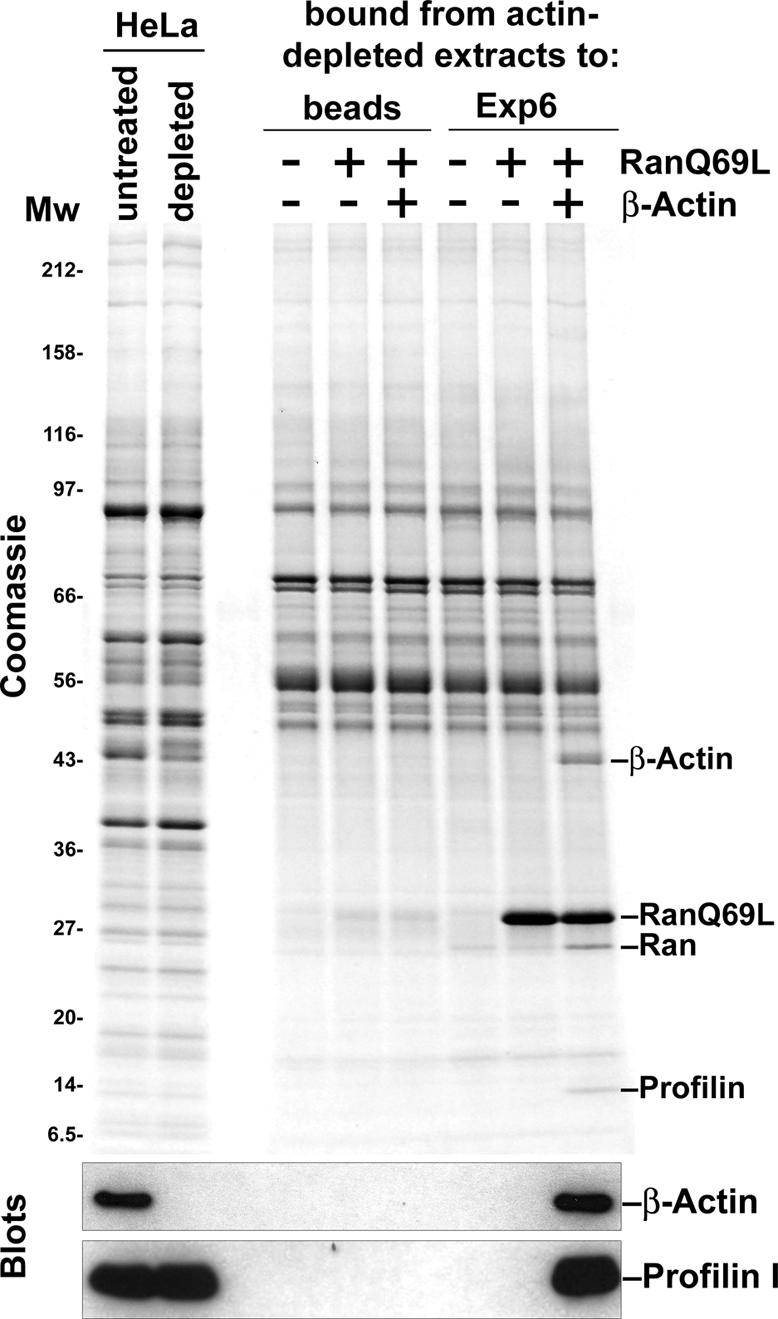
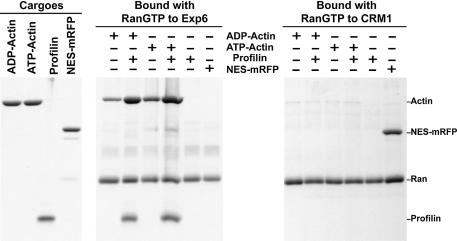
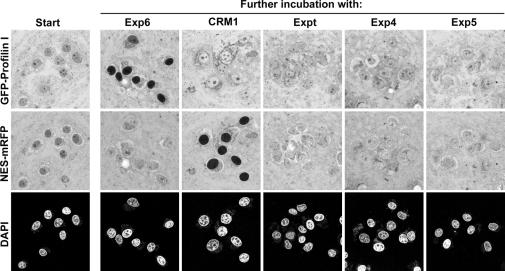
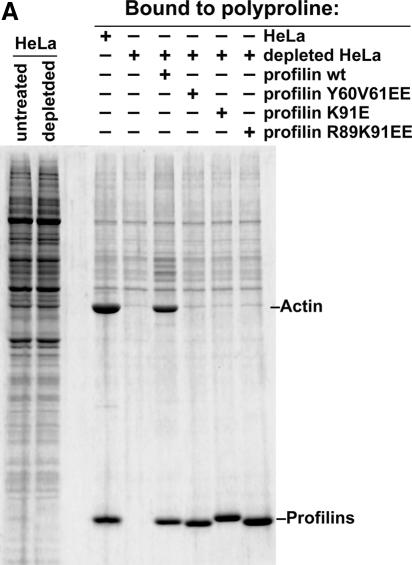
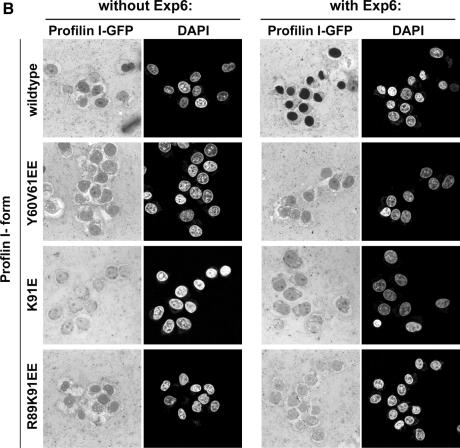
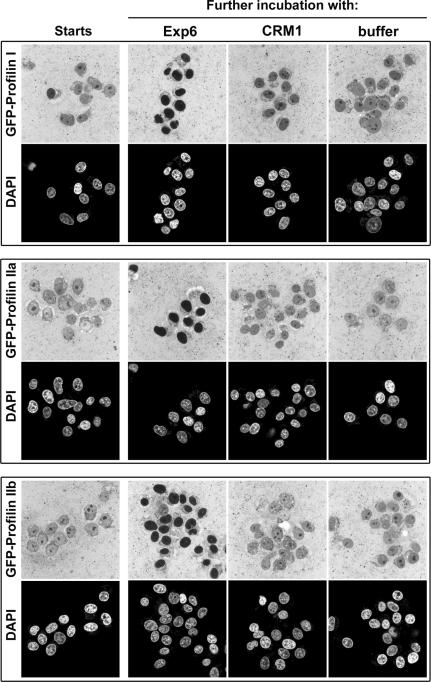
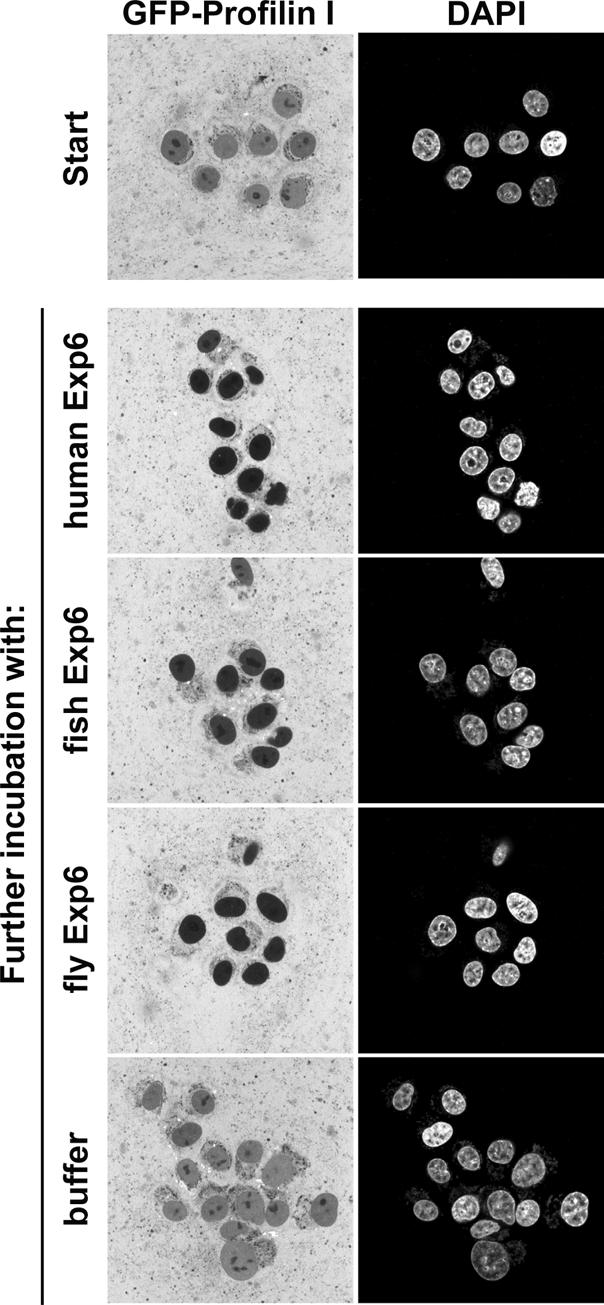
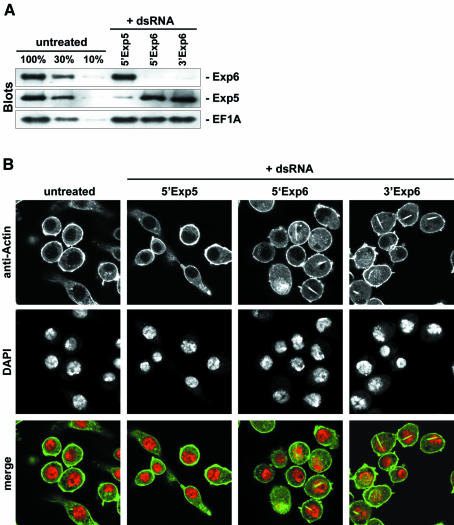
Active macromolecular transport between the nucleus and cytoplasm proceeds through nuclear pore complexes and is mostly mediated by transport receptors of the importin beta-superfamily. Here we identify exportin 6 (Exp6) as a novel family member from higher eukaryotes and show that it mediates nuclear export of profilin.actin complexes. Exp6 appears to contact primarily actin, but the interaction is greatly enhanced by the presence of profilin. Profilin thus functions not only as the nucleotide exchange factor for actin, but can also be regarded as a cofactor of actin export and hence as a suppressor of actin polymerization in the nucleus. Even though human and Drosophila Exp6 share only approximately 20% identical amino acid residues, their function in profilin.actin export is conserved. A knock-down of Drosophila Exp6 by RNA interference abolishes nuclear exclusion of actin and results in the appearance of nuclear actin paracrystals. In contrast to a previous report, we found no indications of a major and direct role for CRM1 in actin export from mammalian or insect nuclei.
Figures
References
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
