

Sequences that direct significant levels of frameshifting are frequent in coding regions of Escherichia coli
- PMID: 14592990
- PMCID: PMC275418
- DOI: 10.1093/emboj/cdg561
Sequences that direct significant levels of frameshifting are frequent in coding regions of Escherichia coli
Abstract
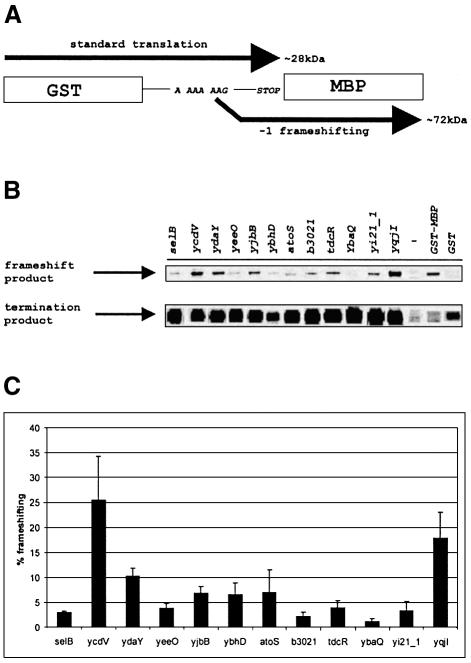
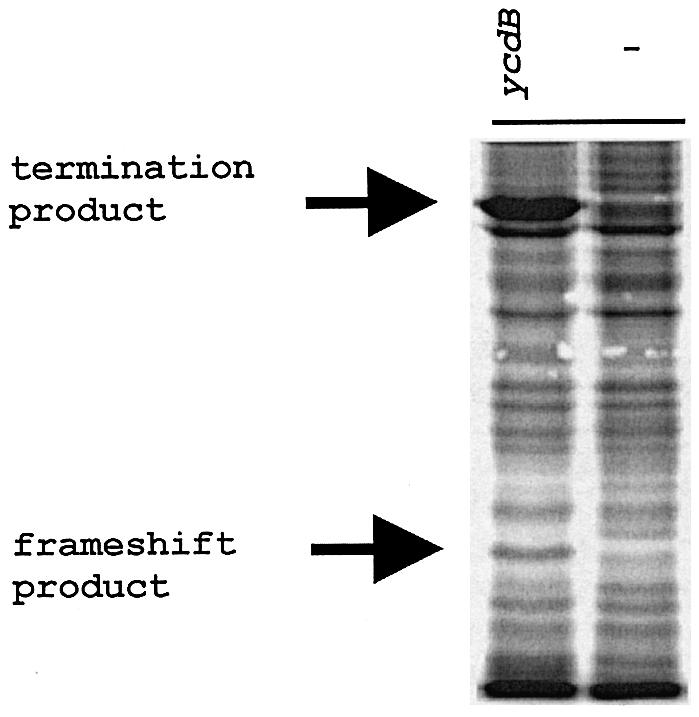
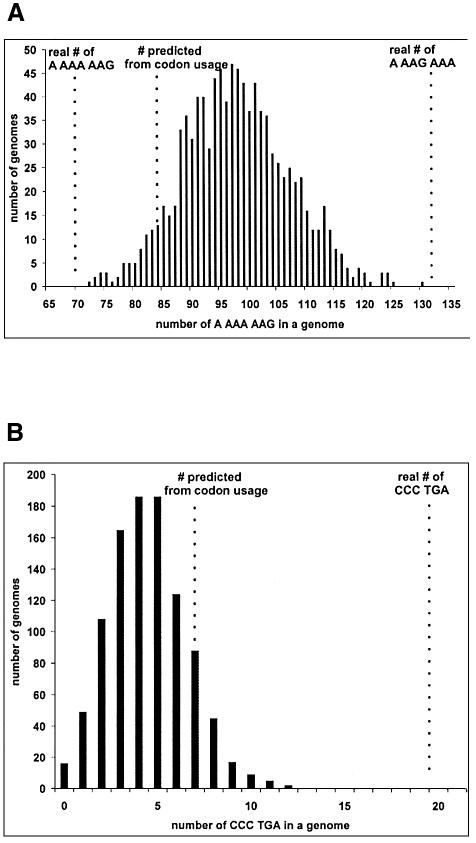
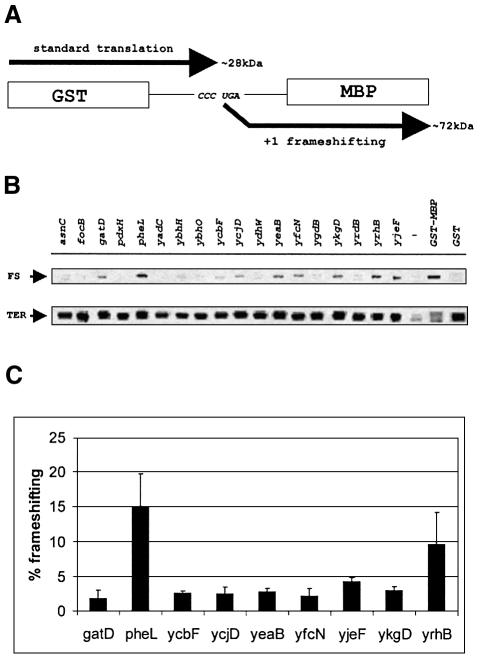
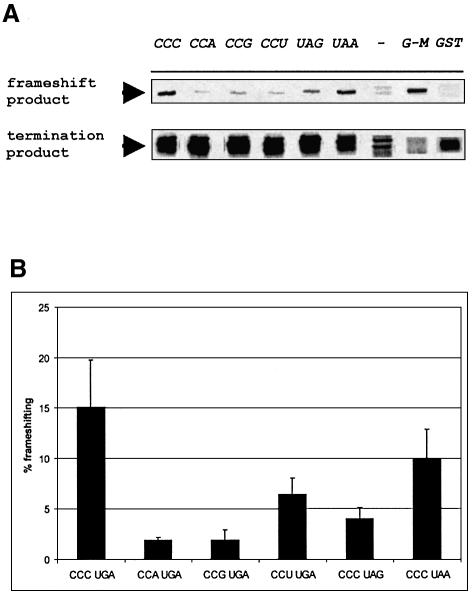
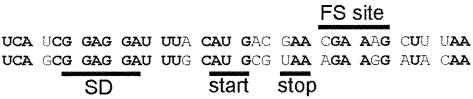
It is generally believed that significant ribosomal frameshifting during translation does not occur without a functional purpose. The distribution of two frameshift-prone sequences, A_AAA_AAG and CCC_TGA, in coding regions of Escherichia coli has been analyzed. Although a moderate level of selection against the first sequence is evident, 68 genes contain A_AAA_AAG and 19 contain CCC_TGA. The majority of those tested in their genomic context showed >1% frameshifting. Comparative sequence analysis was employed to assess a potential biological role for frameshifting in decoding these genes. Two new candidates, in pheL and ydaY, for utilized frameshifting have been identified in addition to those previously known in dnaX and nine insertion sequence elements. For the majority of the shift-prone sequences no functional role can be attributed to them, and the frameshifting is likely erroneous. However, none of frameshift sequences is in the 306 most highly expressed genes. The unexpected conclusion is that moderate frameshifting during expression of at least some other genes is not sufficiently harmful for cells to trigger strong negative evolutionary pressure.
Figures
References
-
- Bang M.L., Centner,T., Fornoff,F., Geach,A.J., Gotthardt,M., McNabb,M., Witt,C.C., Labeit,D., Gregorio,C.C., Granzier,H. and Labeit,S. (2001) The complete gene sequence of titin, expression of an unusual approximately 700-kDa titin isoform, and its interaction with obscurin identify a novel Z-line to I-band linking system. Circ. Res., 89, 1065–1072. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
