

Mss51p promotes mitochondrial Cox1p synthesis and interacts with newly synthesized Cox1p
- PMID: 14592991
- PMCID: PMC275423
- DOI: 10.1093/emboj/cdg566
Mss51p promotes mitochondrial Cox1p synthesis and interacts with newly synthesized Cox1p
Abstract
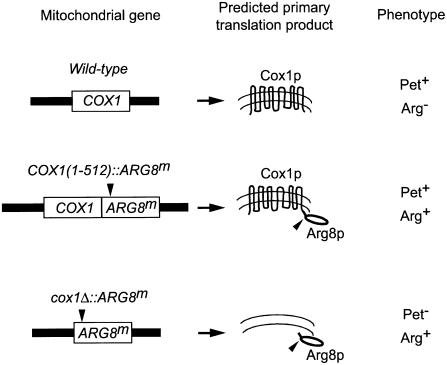
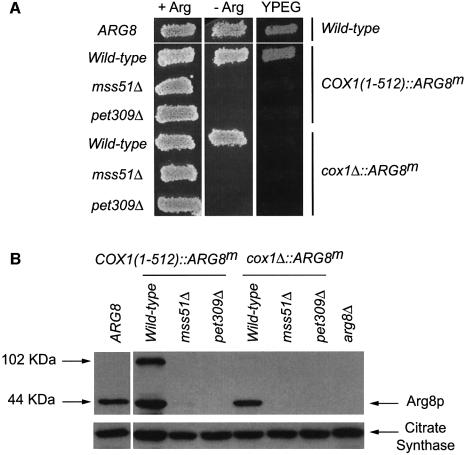
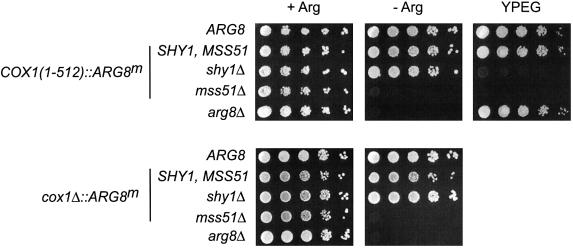
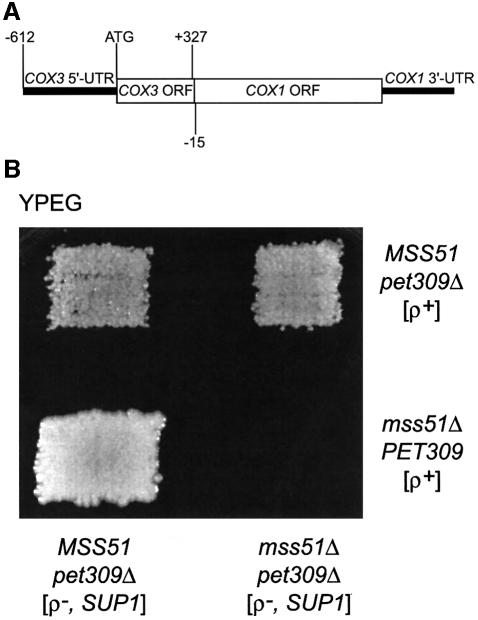
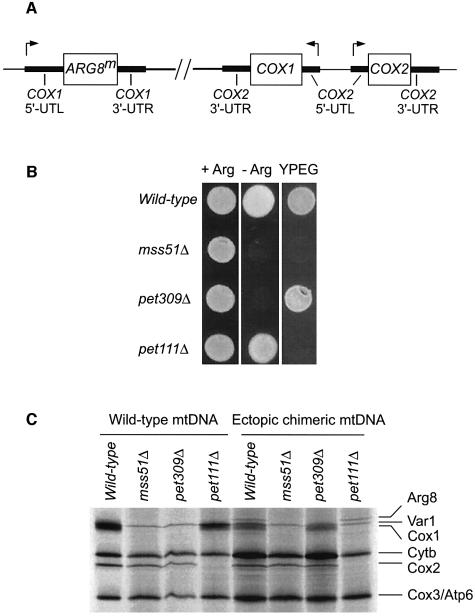
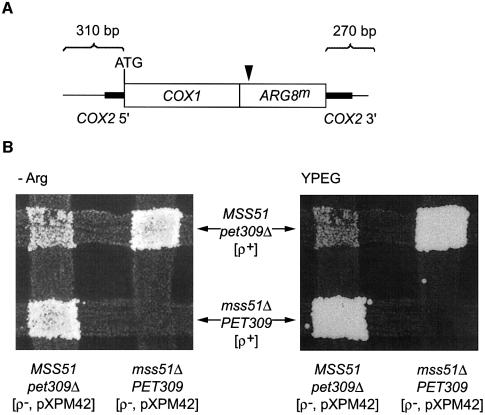
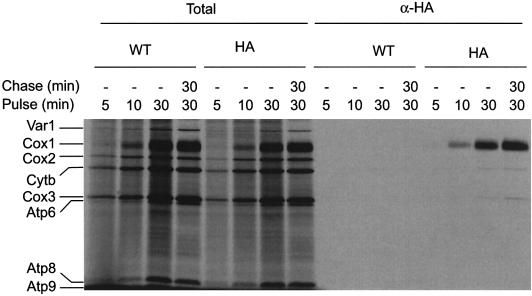
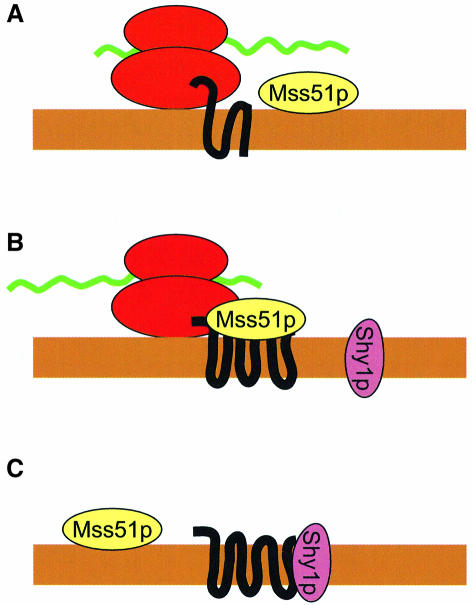
The post-transcriptional role of Mss51p in mitochondrial gene expression is of great interest since MSS51 mutations suppress the respiratory defect caused by shy1 mutations. SHY1 is a Saccharomyces cerevisiae homolog of human SURF1, which when mutated causes a cytochrome oxidase assembly defect. We found that MSS51 is required for expression of the mitochondrial reporter gene ARG8(m) when it is inserted at the COX1 locus, but not when it is at COX2 or COX3. Unlike the COX1 mRNA-specific translational activator PET309, MSS51 has at least two targets in COX1 mRNA. MSS51 acts in the untranslated regions of the COX1 mRNA, since it was required to synthesize Arg8p when ARG8(m) completely replaced the COX1 codons. MSS51 also acts on a target specified by the COX1 coding region, since it was required to translate either COX1 or COX1:: ARG8(m) coding sequences from an ectopic COX2 locus. Mss51p was found to interact physically with newly synthesized Cox1p, suggesting that it could coordinate Cox1p synthesis with insertion into the inner membrane or cytochrome oxidase assembly.
Figures
References
-
- Aldridge P. and Hughes,K.T. (2002) Regulation of flagellar assembly. Curr. Opin. Microbiol., 5, 160–165. - PubMed
-
- Barrientos A., Barros,M.H., Valnot,I., Rotig,A., Rustin,P. and Tzagoloff,A. (2002a) Cytochrome oxidase in health and disease. Gene, 286, 53–63. - PubMed
-
- Bonnefoy N. and Fox,T.D. (2000) In vivo analysis of mutated initiation codons in the mitochondrial COX2 gene of Saccharomyces cerevisiae fused to the reporter gene ARG8m reveals lack of downstream reinitiation. Mol. Gen. Genet., 262, 1036–1046. - PubMed
-
- Bonnefoy N. and Fox,T.D. (2001) Genetic transformation of Saccharomyces cerevisiae mitochondria. Methods Cell Biol., 65, 381–396. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
