

Pulsatile versus oscillatory shear stress regulates NADPH oxidase subunit expression: implication for native LDL oxidation
- PMID: 14593003
- PMCID: PMC4433384
- DOI: 10.1161/01.RES.0000104087.29395.66
Pulsatile versus oscillatory shear stress regulates NADPH oxidase subunit expression: implication for native LDL oxidation
Abstract
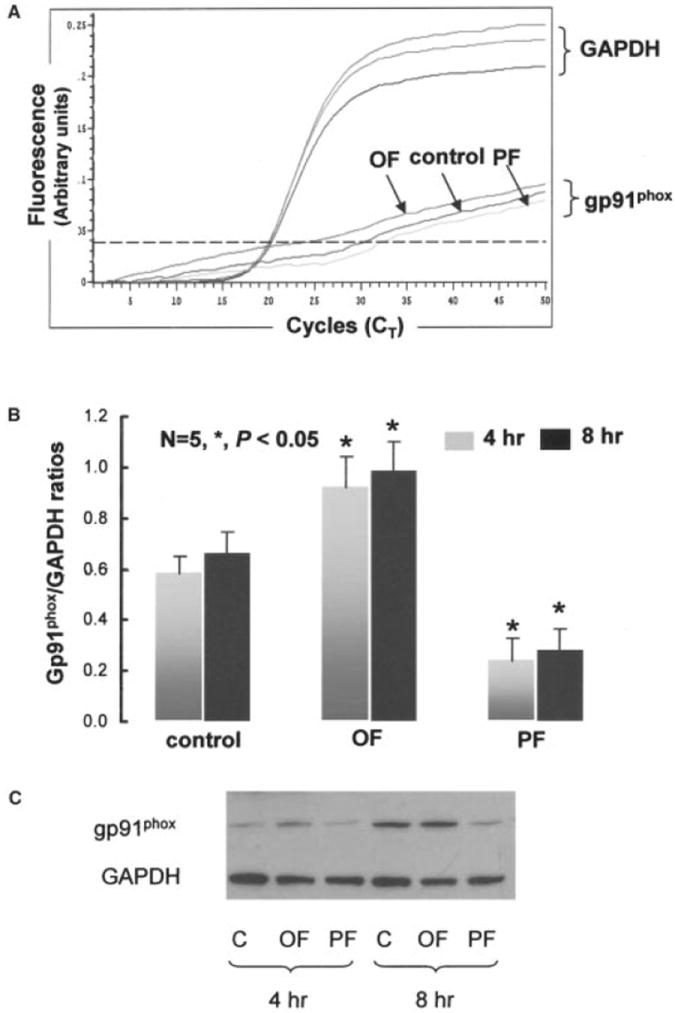
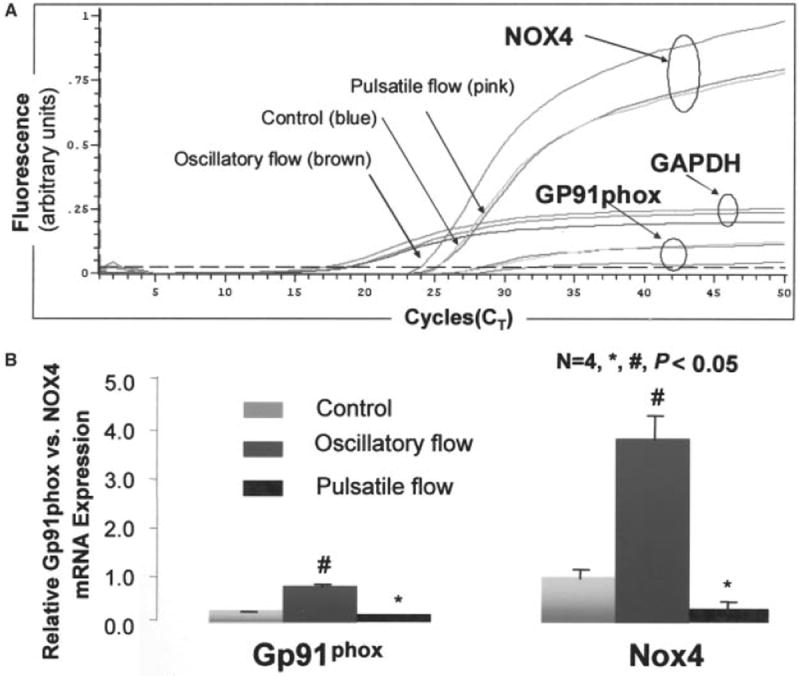
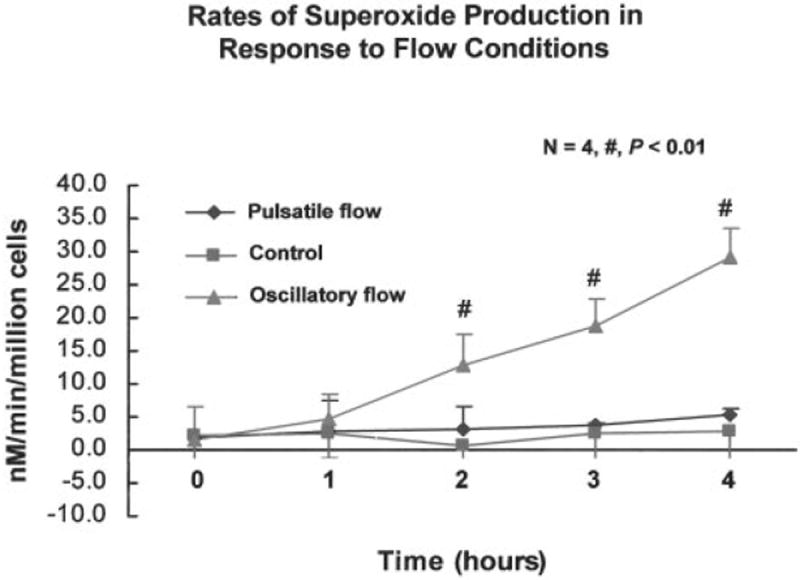
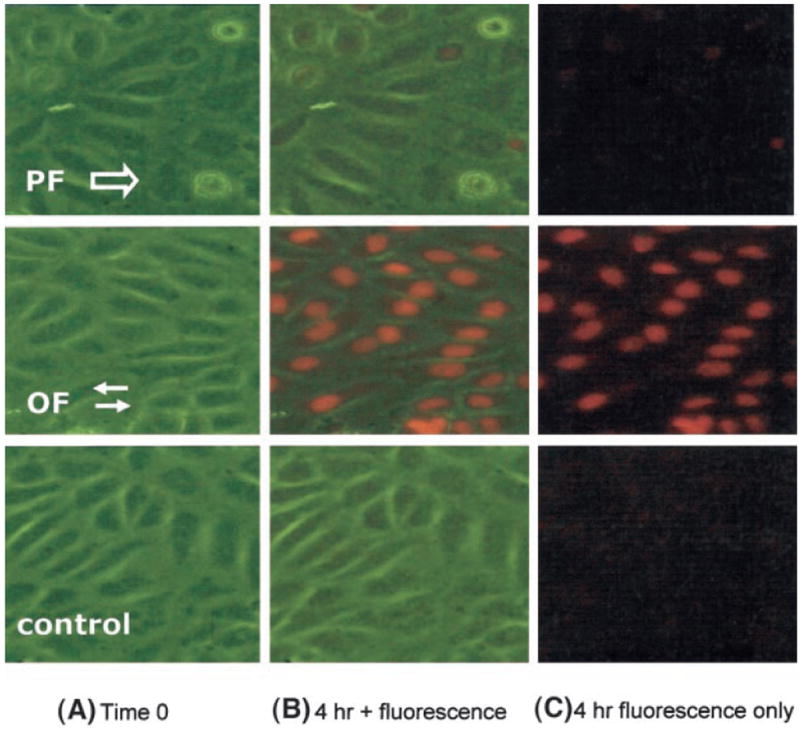
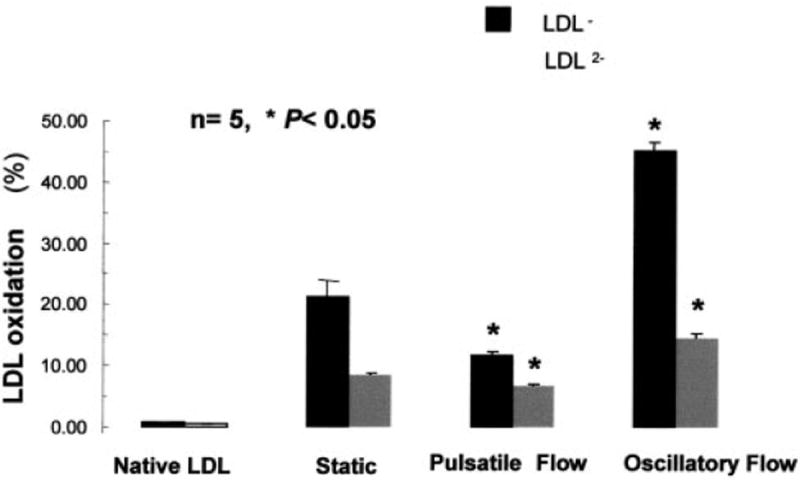
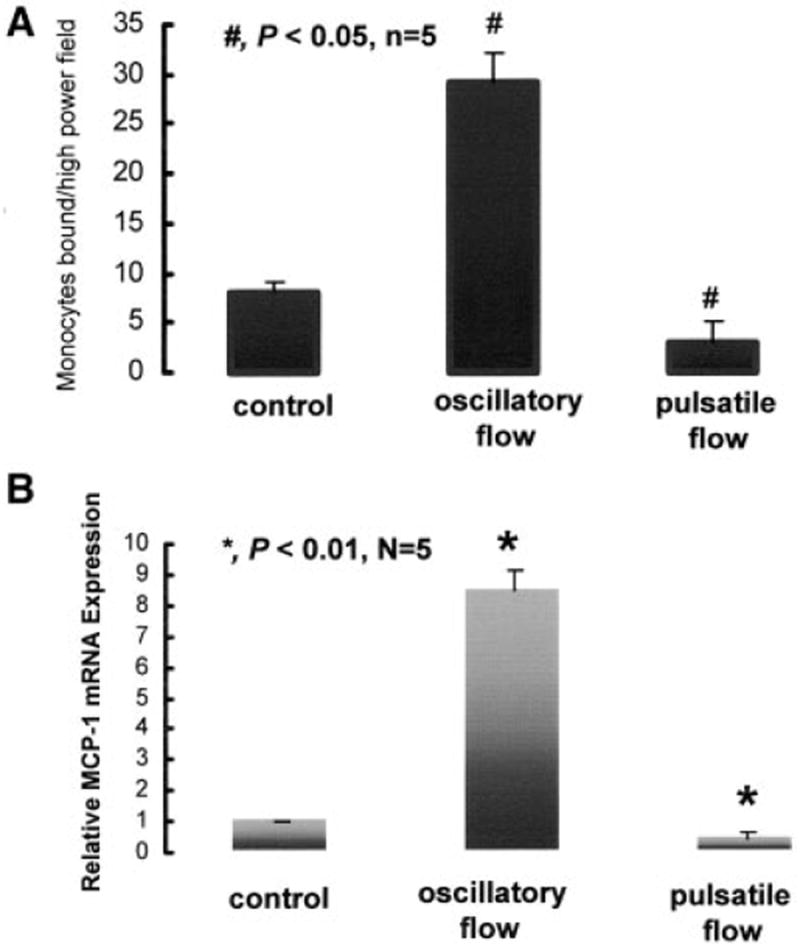
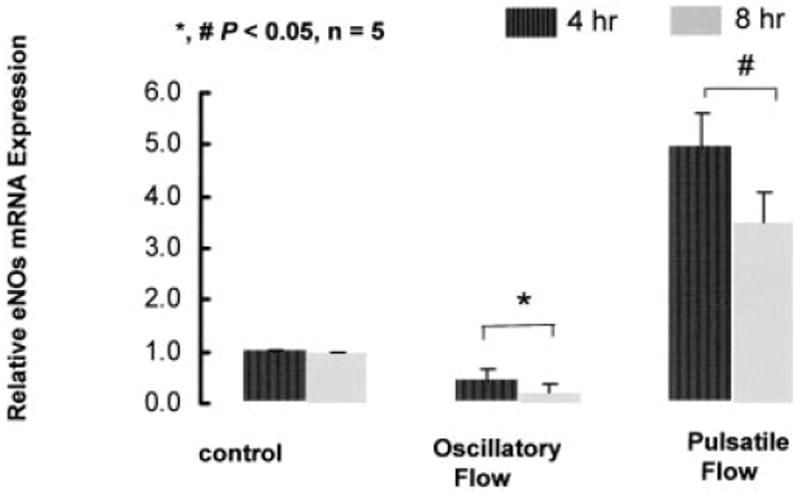
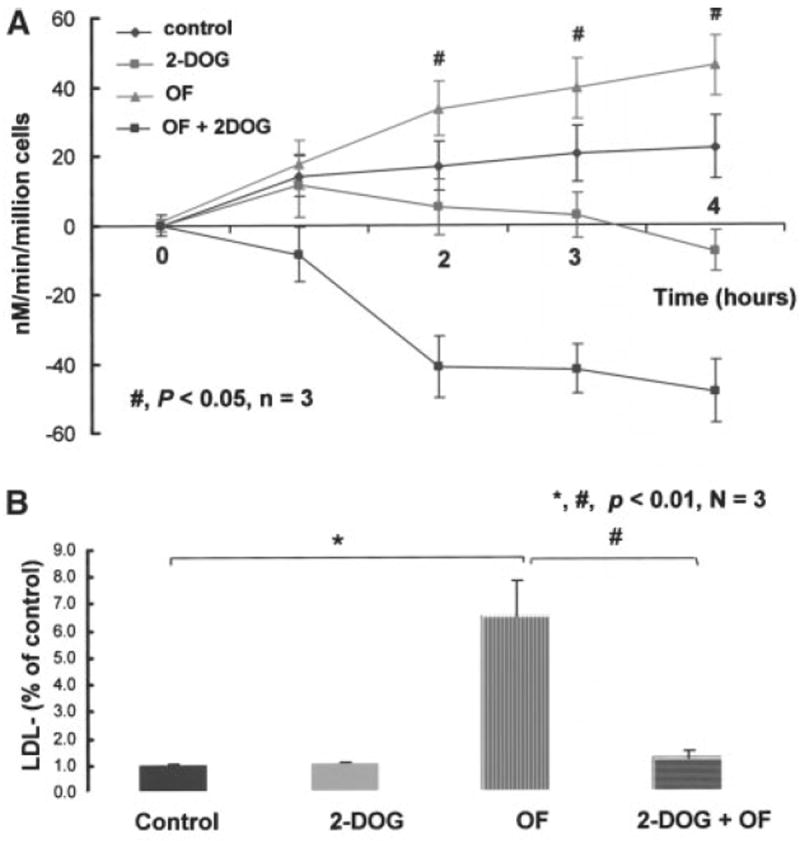
Shear stress regulates endothelial nitric oxide and superoxide (O2-*) production, implicating the role of NADPH oxidase activity. It is unknown whether shear stress regulates the sources of reactive species production, consequent low-density lipoprotein (LDL) modification, and initiation of inflammatory events. Bovine aortic endothelial cells (BAECs) in the presence of 50 microg/mL of native LDL were exposed to (1) pulsatile flow with a mean shear stress (tau(ave)) of 25 dyne/cm2 and (2) oscillating flow at tau(ave) of 0. After 4 hours, aliquots of culture medium were collected for high-performance liquid chromatography analyses of electronegative LDL species, described as LDL- and LDL2-. In response to oscillatory shear stress, gp91phox mRNA expression was upregulated by 2.9+/-0.3-fold, and its homologue, Nox4, by 3.9+/-0.9-fold (P<0.05, n=4), with a corresponding increase in O2-* production rate. The proportion of LDL- and LDL2- relative to static conditions increased by 67+/-17% and 30+/-7%, respectively, with the concomitant upregulation of monocyte chemoattractant protein-1 expression and increase in monocyte/BAEC binding (P<0.05, n=5). In contrast, pulsatile flow downregulated both gp91phox and Nox4 mRNA expression (by 1.8+/-0.2-fold and 3.0+/-0.12-fold, respectively), with an accompanying reduction in O2-* production, reduction in the extent of LDL modification (51+/-12% for LDL- and 30+/-7% for LDL2-), and monocyte/BAEC binding. The flow-dependent LDL oxidation is determined in part by the NADPH oxidase activity. The formation of modified LDL via O2-* production may also affect the regulation of monocyte chemoattractant protein-1 expression and monocyte/BAEC binding.
Figures
Similar articles
-
Pulsatile flow regulates monocyte adhesion to oxidized lipid-induced endothelial cells.Arterioscler Thromb Vasc Biol. 2001 Nov;21(11):1770-6. doi: 10.1161/hq1001.097104. Arterioscler Thromb Vasc Biol. 2001. PMID: 11701464
-
Bone morphogenic protein 4 produced in endothelial cells by oscillatory shear stress induces monocyte adhesion by stimulating reactive oxygen species production from a nox1-based NADPH oxidase.Circ Res. 2004 Oct 15;95(8):773-9. doi: 10.1161/01.RES.0000145728.22878.45. Epub 2004 Sep 23. Circ Res. 2004. PMID: 15388638
-
NO-mediated regulation of NAD(P)H oxidase by laminar shear stress in human endothelial cells.J Physiol. 2006 Oct 15;576(Pt 2):557-67. doi: 10.1113/jphysiol.2006.111070. Epub 2006 Jul 27. J Physiol. 2006. PMID: 16873416 Free PMC article.
-
Mechanosensitive microRNAs-role in endothelial responses to shear stress and redox state.Free Radic Biol Med. 2013 Sep;64:61-8. doi: 10.1016/j.freeradbiomed.2013.05.034. Epub 2013 May 30. Free Radic Biol Med. 2013. PMID: 23727269 Free PMC article. Review.
-
A central role for the endothelial NADPH oxidase in atherosclerosis.FEBS Lett. 2000 Apr 21;472(1):1-4. doi: 10.1016/s0014-5793(00)01397-1. FEBS Lett. 2000. PMID: 10781793 Review.
Cited by
-
Deletion of NoxO1 limits atherosclerosis development in female mice.Redox Biol. 2020 Oct;37:101713. doi: 10.1016/j.redox.2020.101713. Epub 2020 Sep 4. Redox Biol. 2020. PMID: 32949971 Free PMC article.
-
Nephroprotection of lacidipine against gentamycin-induced nephrotoxicity in albino rats.J Exp Pharmacol. 2010 Jun 3;2:59-63. doi: 10.2147/jep.s10978. eCollection 2010. J Exp Pharmacol. 2010. PMID: 27186092 Free PMC article.
-
LDL phospholipid hydrolysis produces modified electronegative particles with an unfolded apoB-100 protein.J Lipid Res. 2005 Jan;46(1):115-22. doi: 10.1194/jlr.M400306-JLR200. Epub 2004 Oct 16. J Lipid Res. 2005. PMID: 15489541 Free PMC article.
-
Hemodynamics of distal cerebral arteries are associated with functional outcomes in symptomatic ischemic stroke in middle cerebral artery territory: A four-dimensional flow cardiovascular magnetic resonance study.J Cardiovasc Magn Reson. 2025 Summer;27(1):101857. doi: 10.1016/j.jocmr.2025.101857. Epub 2025 Feb 10. J Cardiovasc Magn Reson. 2025. PMID: 39938618 Free PMC article.
-
Hydrogen peroxide induces nitric oxide and proteosome activity in endothelial cells: a bell-shaped signaling response.Free Radic Biol Med. 2007 Apr 1;42(7):1049-61. doi: 10.1016/j.freeradbiomed.2007.01.005. Epub 2007 Jan 8. Free Radic Biol Med. 2007. PMID: 17349932 Free PMC article.
References
-
- Dzau VJ. Theodore Cooper Lecture. Tissue angiotensin and pathobiology of vascular disease: a unifying hypothesis. Hypertension. 2001;37:1047–1052. - PubMed
-
- Hsieh HJ, Cheng CC, Wu ST, Chiu JJ, Wung BS, Wang DL. Increase of reactive oxygen species (ROS) in endothelial cells by shear flow and involvement of ROS in shear-induced c-fos expression. J Cell Physiol. 1998;175:156–162. - PubMed
-
- Mollnau H, Wendt M, Szocs K, Lassegue B, Schulz E, Oelze M, Li H, Bodenschatz M, August M, Kleschyov AL, Tsilimingas N, Walter U, Forstermann U, Meinertz T, Griendling K, Munzel T. Effects of angiotensin II infusion on the expression and function of NAD(P)H oxidase and components of nitric oxide/cGMP signaling. Circ Res. 2002;90:e58–e65. - PubMed
-
- Nickenig G, Harrison DG. The AT1-type angiotensin receptor in oxidative stress and atherogenesis, part I: oxidative stress and atherogenesis. Circulation. 2002;105:393–396. - PubMed
-
- Sowers JR, Epstein M, Frohlich ED. Diabetes, hypertension, and cardiovascular disease: an update. Hypertension. 2001;37:1053–1059. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
