

doi: 10.1128/JB.185.22.6719-6722.2003.
Translocon "pulling" of nascent SecM controls the duration of its translational pause and secretion-responsive secA regulation
Affiliations
- PMID: 14594848
- PMCID: PMC262105
- DOI: 10.1128/JB.185.22.6719-6722.2003
Item in Clipboard
Translocon "pulling" of nascent SecM controls the duration of its translational pause and secretion-responsive secA regulation
J Bacteriol.
2003 Nov.
Abstract
SecA is an ATPase and motor protein that drives protein translocation across the bacterial plasma membrane. In Escherichia coli SecA levels are regulated by the secretion needs of the cell utilizing secM, which encodes a secreted protein. Previous studies demonstrated that this regulation requires a translational pause within secM, whose duration regulates the accessibility of the secA Shine-Dalgarno sequence on secM secA mRNA. Here we provide evidence that translocon "pulling" of nascent SecM is what regulates the duration of the secM translational pause, and thus secA expression levels, thereby providing direct support for this model.
Figures
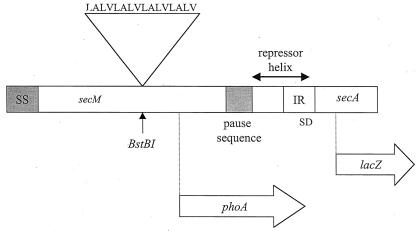
Creation of an artificial membrane anchor sequence in secM. The membrane anchor peptide sequence is shown along with a diagram of the secM secA operon that indicates the location of the atypical secM signal (SS) and translational pause sequences (codons 1 to 37 and 150 to 166, respectively) and the repressor helix that blocks the secA Shine-Dalgarno sequence (SD) (12, 22). The sites of insertion of the membrane anchor sequence within secM (BstBI at codon 74), the secM-phoA fusion joint (codon 157), and the secA-lacZ fusion joint (codon 67) are indicated. Only the 5′ portion of secA is shown.
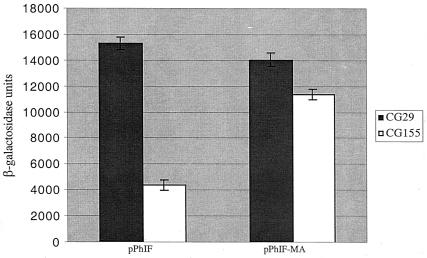
Effect of the secM membrane anchor sequence on secA regulation. CG155 (MC1000 recA) and CG29 [MC1000 secD1(Cs) phoR recA1 srl::Tn10] containing pPhIF or pPhIF-MA were grown in Luria broth containing 100 μg of ampicillin per ml at 39°C to mid-logarithmic phase when the cultures were shifted to 23°C for 4 h. β-Galactosidase assays were performed in duplicate for each of two duplicate cultures as described previously (11). The average result is given with the error bar indicating the standard deviation.
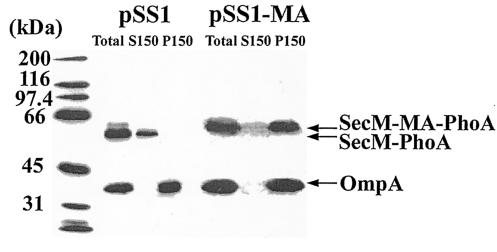
Subcellular location of the SecM-PhoA chimeras. CC118(pSS1) and CC118(pSS1-MA) were grown in Luria broth containing 100 μg of ampicillin per ml at 30°C to an optical density at 600 nm of 1.0, when cells were harvested by sedimentation at 7,000 × g for 5 min at 4°C. Cell pellets were resuspended in 0.02 volumes of TKMDP (25 mM TrisOAc [pH 7.5], 25 mM KCl, 1 mM MgOAc, 1 mM dithiothreitol, 0.5 mM EDTA, 0.5 mM phenylmethylsulfonyl fluoride, 100 μM leupeptin, 1 μM pepstatin, and 0.3 μM aprotinin) and broken by two passages at 8,000 lb/in2 in the French pressure cell. Unbroken cells were removed by two successive sedimentations at 13,000 × g for 10 min at 4°C, giving rise to the total cleared lysate (Total). Soluble (S150) and membrane (P150) fractions were obtained by sedimentation at 150,000 × g for 3 h at 4°C. The P150 fraction was resuspended in the original volume of TKMDP. A 28-μl [CC118(pSS1)] or 56-μl [CC118(pSS1-MA)] quantity of each fraction was analyzed by sodium dodecyl sulfate-polyacrylamide gel electrophoresis followed by Western blotting with alkaline phosphatase and OmpA antiserum and visualization with enhanced chemiluminescence as described by the manufacturer (Perkin-Elmer). The positions of the wild type (SecM-PhoA) and the SecM-PhoA chimera containing the membrane anchor (SecM-MA-PhoA) are given along with that of OmpA. Molecular mass markers are provided at the left.
References
-
- Danese, P. N., and T. J. Silhavy. 1998. Targeting and assembly of periplasmic and outer-membrane proteins in Escherichia coli. Annu. Rev. Genet. 32:59-94. - PubMed
-
- Davis, N. G., and P. Model. 1985. An artificial anchor domain: hydrophobicity suffices to stop transfer. Cell 41:607-614. - PubMed
-
- Economou, A., and W. Wickner. 1994. SecA promotes preprotein translocation by undergoing ATP-driven cycles of membrane insertion and deinsertion. Cell 78:835-843. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
