

On the role of the two extracytoplasmic substrate-binding domains in the ABC transporter OpuA
- PMID: 14609945
- PMCID: PMC275439
- DOI: 10.1093/emboj/cdg581
On the role of the two extracytoplasmic substrate-binding domains in the ABC transporter OpuA
Abstract
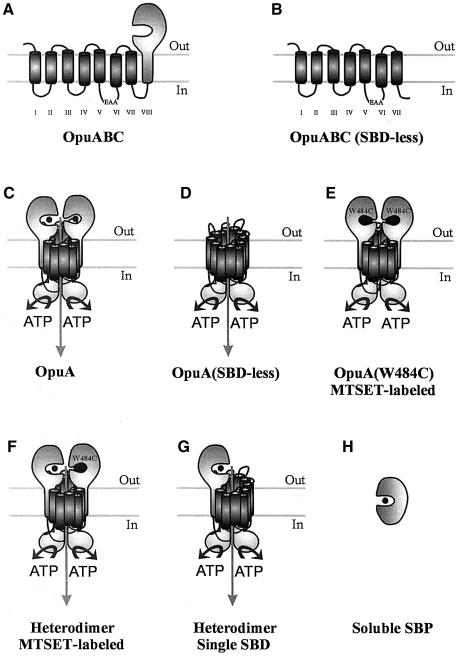
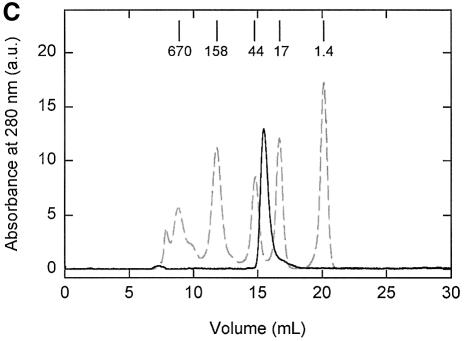
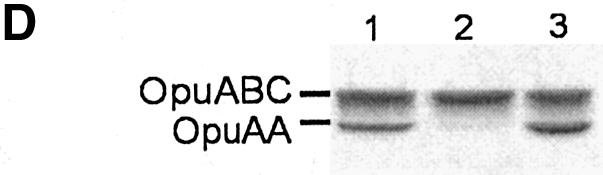
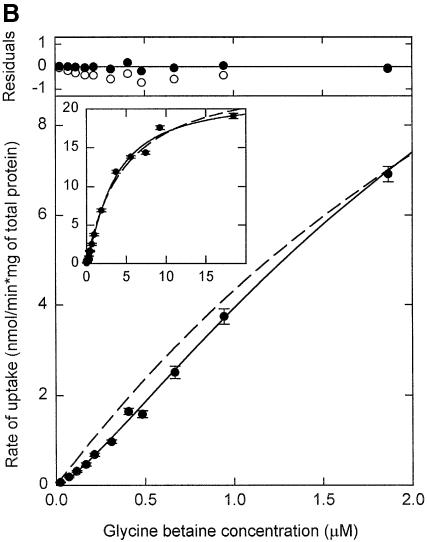
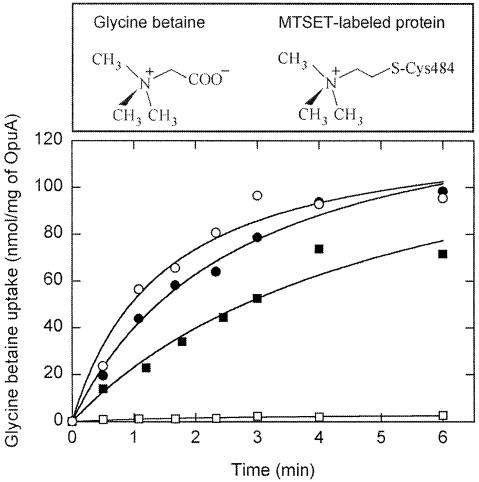
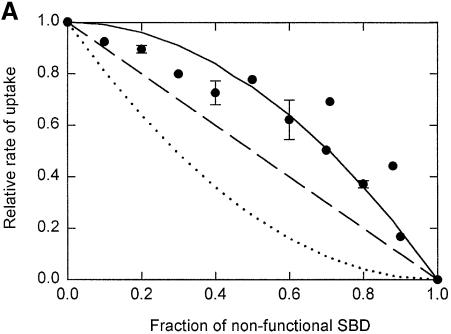
Members of two transporter families of the ATP-binding cassette (ABC) superfamily use two or even four extracytoplasmic substrate-binding domains (SBDs) for transport. We report on the role of the two SBDs in the translocation cycle of the ABC transporter OpuA from Lactococcus lactis. Heterooligomeric OpuA complexes with only one SBD or one functional and one non-functional SBD (inactivated by covalent linkage of a substrate mimic) have been constructed, and the substrate binding and transport kinetics of the purified transporters, reconstituted in liposomes, have been determined. The data indicate that the two SBDs of OpuA interact in a cooperative manner in the translocation process by stimulating either the docking of the SBDs onto the translocator or the delivery of glycine betaine to the translocator. It appears that one of these initial steps, but not the later steps in translocation or resetting of the system to the initial state, is rate determining for transport. These new insights on the functional role of the extracytoplasmic SBDs are discussed in the light of the current knowledge of substrate-binding-protein-dependent ABC transporters.
Figures







References
-
- Ames G.F., Liu,C.E., Joshi,A.K. and Nikaido,K. (1996) Liganded and unliganded receptors interact with equal affinity with the membrane complex of periplasmic permeases, a subfamily of traffic ATPases. J. Biol. Chem., 271, 14264–14270. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
