

Eme1 is involved in DNA damage processing and maintenance of genomic stability in mammalian cells
- PMID: 14609959
- PMCID: PMC275438
- DOI: 10.1093/emboj/cdg580
Eme1 is involved in DNA damage processing and maintenance of genomic stability in mammalian cells
Abstract
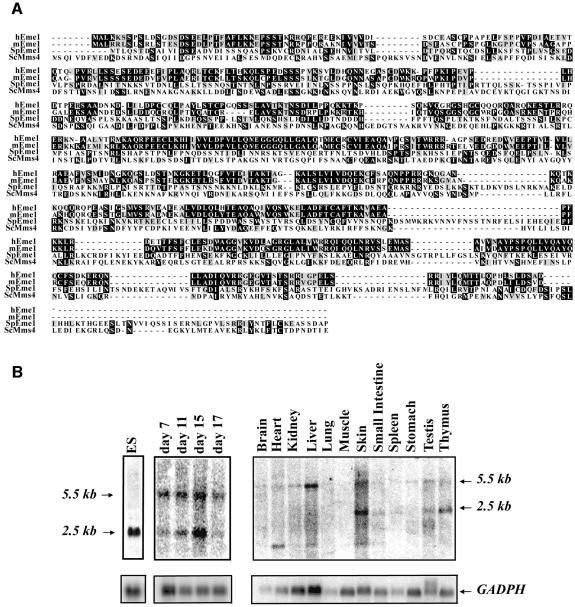
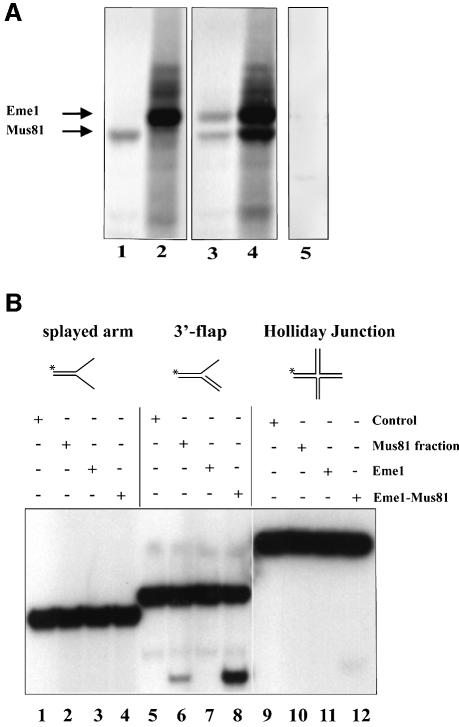
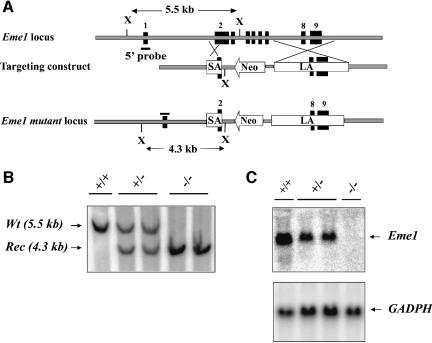
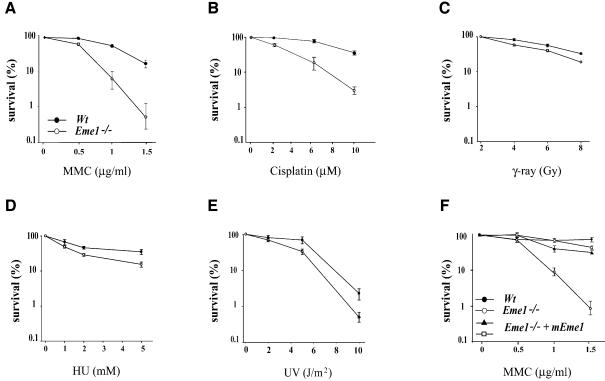
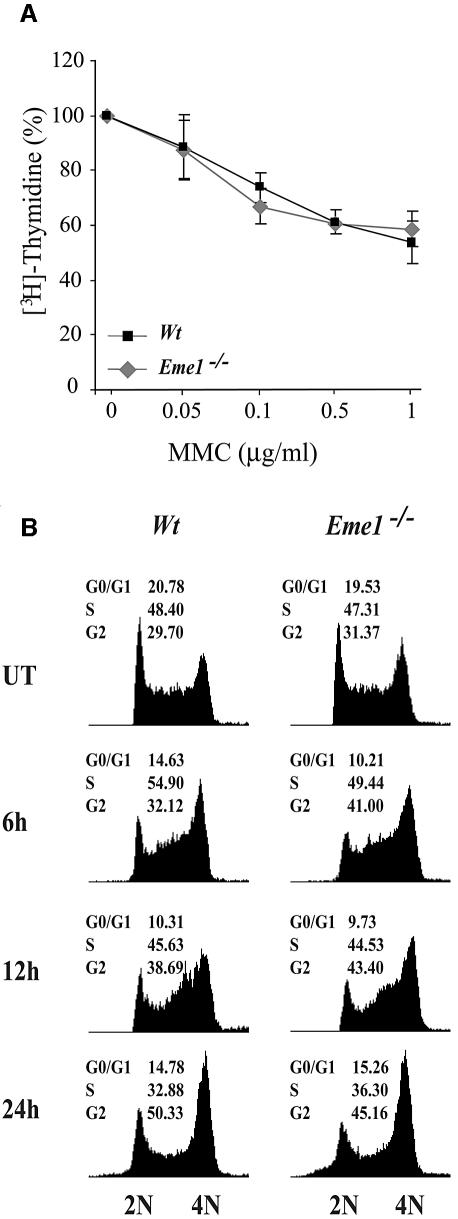
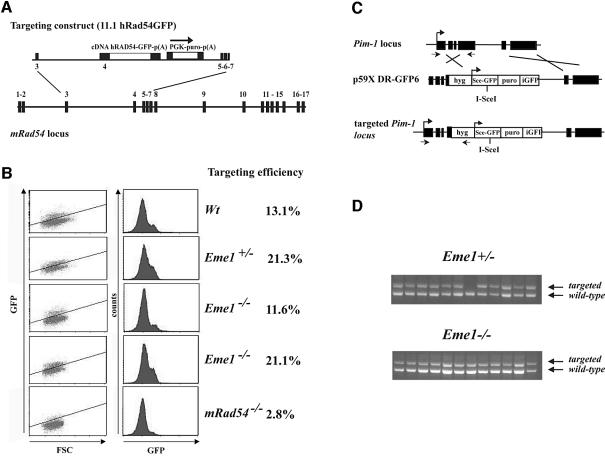
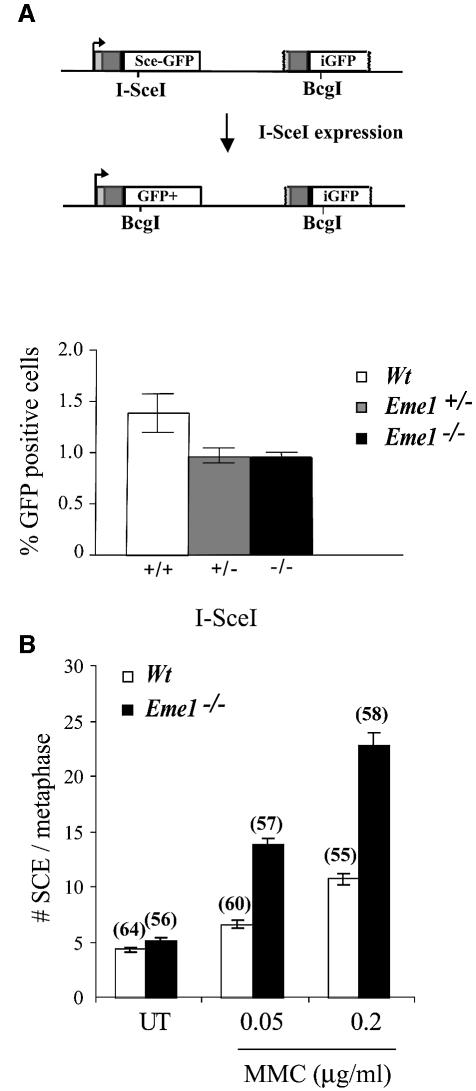
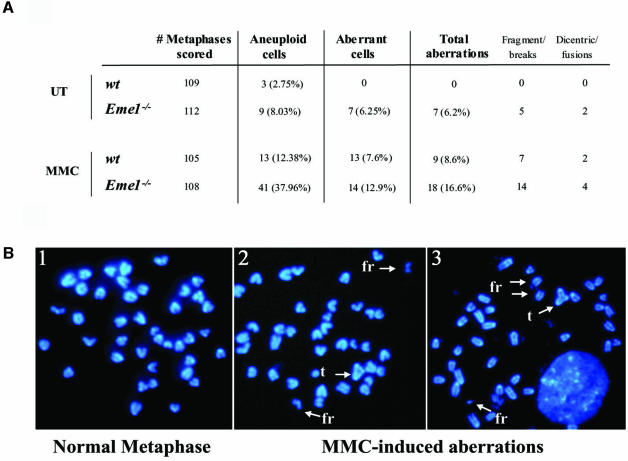
Yeast and human Eme1 protein, in complex with Mus81, constitute an endonuclease that cleaves branched DNA structures, especially those arising during stalled DNA replication. We identified mouse Eme1, and show that it interacts with Mus81 to form a complex that preferentially cleaves 3'-flap structures and replication forks rather than Holliday junctions in vitro. We demonstrate that Eme1-/- embryonic stem (ES) cells are hypersensitive to the DNA cross-linking agents mitomycin C and cisplatin, but only mildly sensitive to ionizing radiation, UV radiation and hydroxyurea treatment. Mammalian Eme1 is not required for the resolution of DNA intermediates that arise during homologous recombination processes such as gene targeting, gene conversion and sister chromatid exchange (SCE). Unlike Blm-deficient ES cells, increased SCE was seen only following induced DNA damage in Eme1-deficient cells. Most importantly, Eme1 deficiency led to spontaneous genomic instability. These results reveal that mammalian Eme1 plays a key role in DNA repair and the maintenance of genome integrity.
Figures
References
-
- Bassing C.H., Swat,W. and Alt,F.W. (2002) The mechanism and regulation of chromosomal V(D)J recombination. Cell, 109, S45–S55. - PubMed
-
- Benson F.E. and West,S.C. (1994) Substrate specificity of the Escherichia coli RuvC protein. Resolution of three- and four-stranded recombination intermediates. J. Biol. Chem., 269, 5195–5201. - PubMed
-
- Boddy M.N., Gaillard,P.H., McDonald,W.H., Shanahan,P., Yates,J.R.,3rd and Russell,P. (2001) Mus81–Eme1 are essential components of a Holliday junction resolvase. Cell, 107, 537–548. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
