

Cloning of the first sn1-DAG lipases points to the spatial and temporal regulation of endocannabinoid signaling in the brain
- PMID: 14610053
- PMCID: PMC2173631
- DOI: 10.1083/jcb.200305129
Cloning of the first sn1-DAG lipases points to the spatial and temporal regulation of endocannabinoid signaling in the brain
Abstract
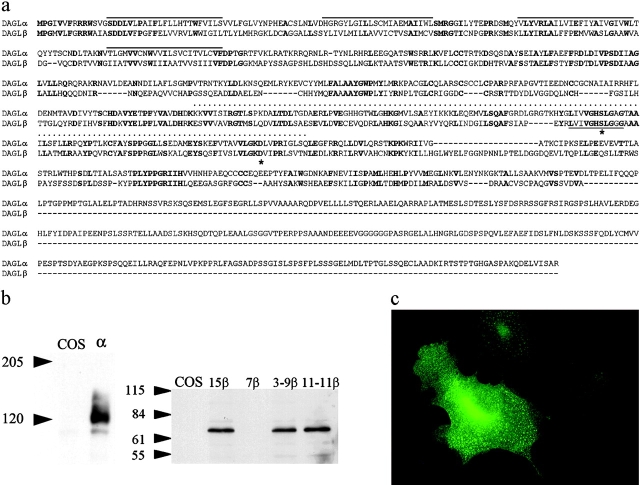
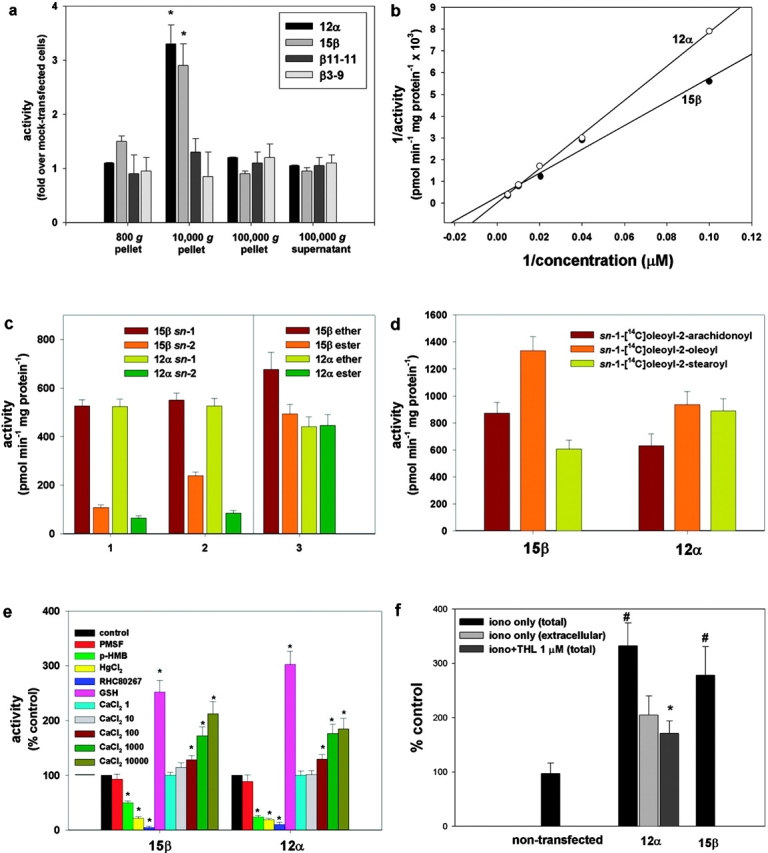
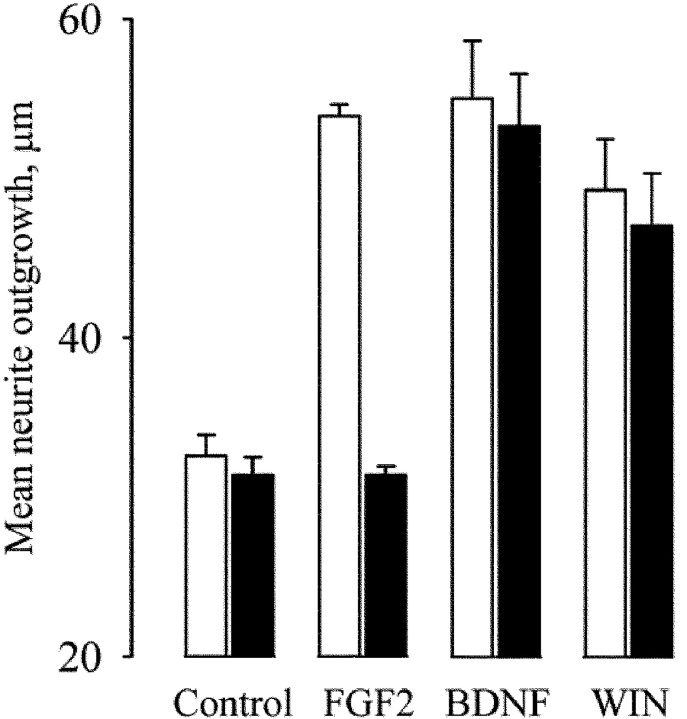
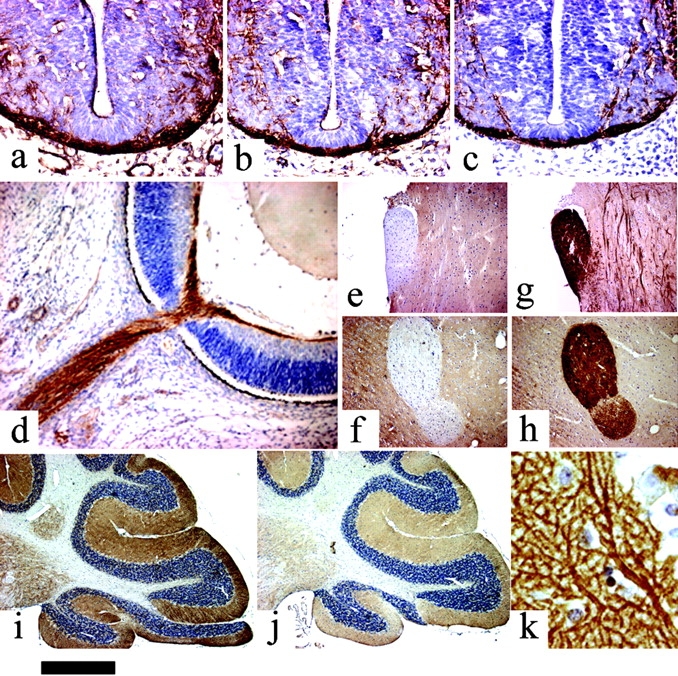
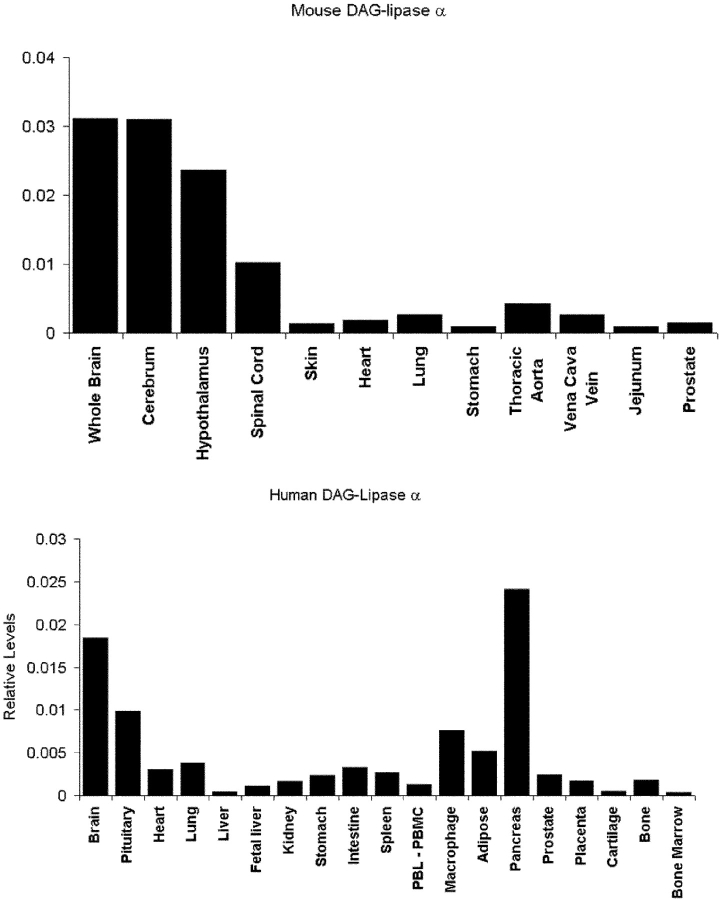
Diacylglycerol (DAG) lipase activity is required for axonal growth during development and for retrograde synaptic signaling at mature synapses. This enzyme synthesizes the endocannabinoid 2-arachidonoyl-glycerol (2-AG), and the CB1 cannabinoid receptor is also required for the above responses. We now report on the cloning and enzymatic characterization of the first specific sn-1 DAG lipases. Two closely related genes have been identified and their expression in cells correlated with 2-AG biosynthesis and release. The expression of both enzymes changes from axonal tracts in the embryo to dendritic fields in the adult, and this correlates with the developmental change in requirement for 2-AG synthesis from the pre- to the postsynaptic compartment. This switch provides a possible explanation for a fundamental change in endocannabinoid function during brain development. Identification of these enzymes may offer new therapeutic opportunities for a wide range of disorders.
Figures
References
-
- Bond, B.C., D.J. Virley, N.J. Cairns, A.J. Hunter, G.B. Moore, S.J. Moss, A.W. Mudge, F.S. Walsh, E. Jazin, and P. Preece. 2002. The quantification of gene expression in an animal model of brain ischaemia using TaqMan real-time RT-PCR. Brain Res. Mol. Brain Res. 106:101–116. - PubMed
-
- Brittis, P.A., J. Silver, F.S. Walsh, and P. Doherty. 1996. Fibroblast growth factor receptor function is required for the orderly projection of ganglion cell axons in the developing mammalian retina. Mol. Cell. Neurosci. 8:120–128. - PubMed
-
- Di Marzo, V., D. Melck, T. Bisogno, and L. De Petrocellis. 1998. Endocannabinoids: endogenous cannabinoid receptor ligands with neuromodulatory action. Trends Neurosci. 21:521–528. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
