

p120 catenin associates with kinesin and facilitates the transport of cadherin-catenin complexes to intercellular junctions
- PMID: 14610057
- PMCID: PMC2173663
- DOI: 10.1083/jcb.200305137
p120 catenin associates with kinesin and facilitates the transport of cadherin-catenin complexes to intercellular junctions
Abstract
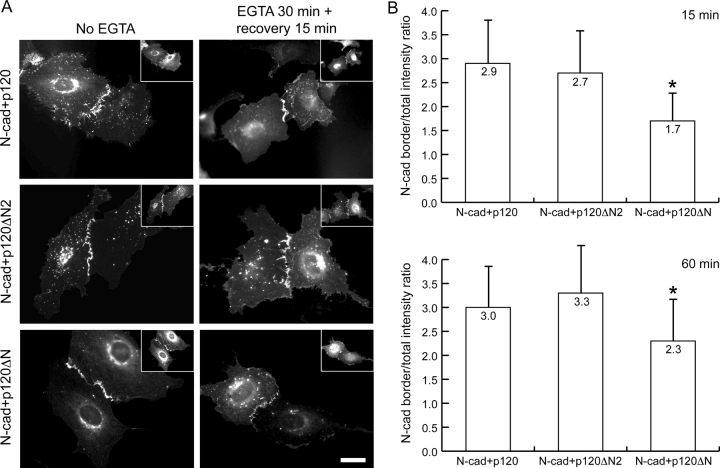
p120 catenin (p120) is a component of adherens junctions and has been implicated in regulating cadherin-based cell adhesion as well as the activity of Rho small GTPases, but its exact roles in cell-cell adhesion are unclear. Using time-lapse imaging, we show that p120-GFP associates with vesicles and exhibits unidirectional movements along microtubules. Furthermore, p120 forms a complex with kinesin heavy chain through the p120 NH2-terminal head domain. Overexpression of p120, but not an NH2-terminal deletion mutant deficient in kinesin binding, recruits endogenous kinesin to N-cadherin. Disruption of the interaction between N-cadherin and p120, or the interaction between p120 and kinesin, leads to a delayed accumulation of N-cadherin at cell-cell contacts during calcium-initiated junction reassembly. Our analyses identify a novel role of p120 in promoting cell surface trafficking of cadherins via association and recruitment of kinesin.
Figures
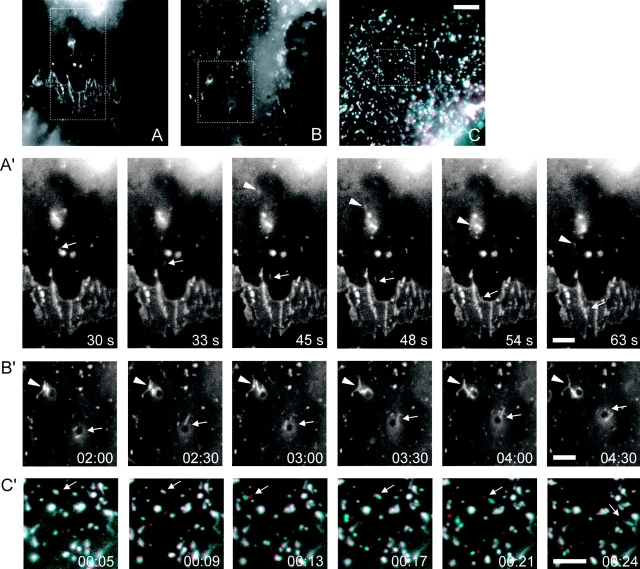
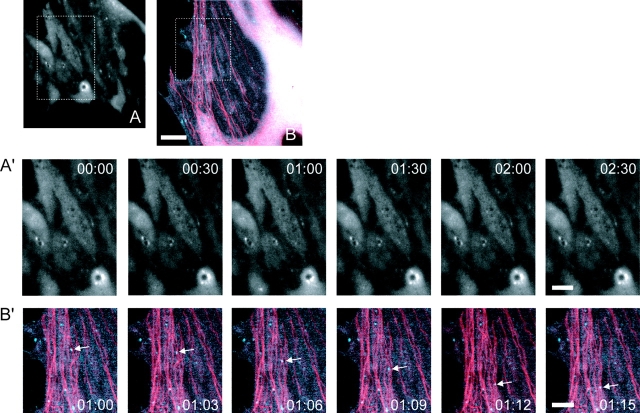

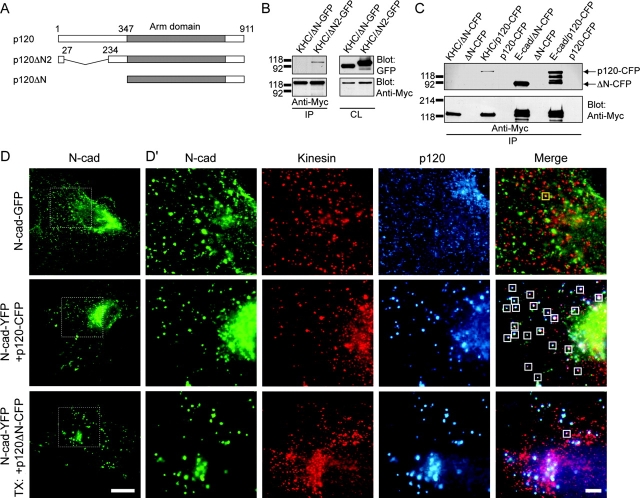

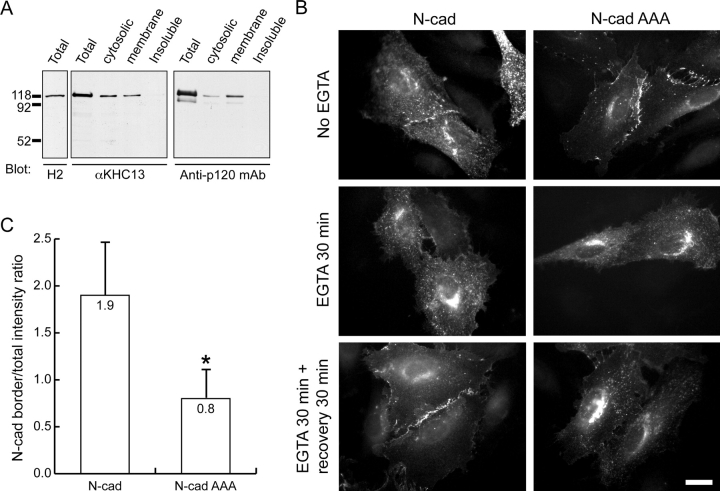
Comment in
-
Traffic control: p120-catenin acts as a gatekeeper to control the fate of classical cadherins in mammalian cells.J Cell Biol. 2003 Nov 10;163(3):437-40. doi: 10.1083/jcb.200310090. J Cell Biol. 2003. PMID: 14610049 Free PMC article. Review.
References
-
- Aberle, H., S. Butz, J. Stappert, H. Weissig, R. Kemler, and H. Hoschuetzky. 1994. Assembly of the cadherin-catenin complex in vitro with recombinant proteins. J. Cell Sci. 107:3655–3663. - PubMed
-
- Anastasiadis, P.Z., and A.B. Reynolds. 2000. The p120 catenin family: complex roles in adhesion, signaling and cancer. J. Cell Sci. 113:1319–1334. - PubMed
-
- Anastasiadis, P.Z., S.Y. Moon, M.A. Thoreson, D.J. Mariner, H.C. Crawford, Y. Zheng, and A.B. Reynolds. 2000. Inhibition of RhoA by p120 catenin. Nat. Cell Biol. 2:637–644. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
