

ROCK-generated contractility regulates breast epithelial cell differentiation in response to the physical properties of a three-dimensional collagen matrix
- PMID: 14610060
- PMCID: PMC2173660
- DOI: 10.1083/jcb.200305010
ROCK-generated contractility regulates breast epithelial cell differentiation in response to the physical properties of a three-dimensional collagen matrix
Abstract
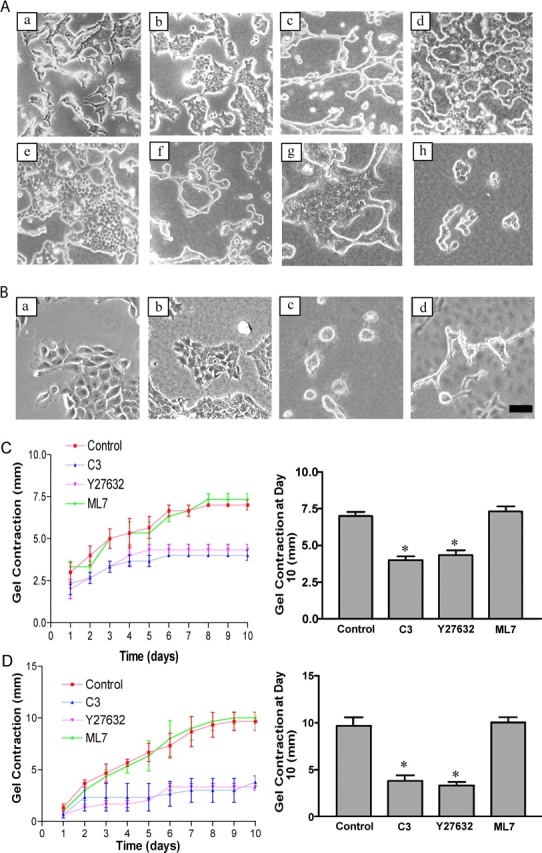
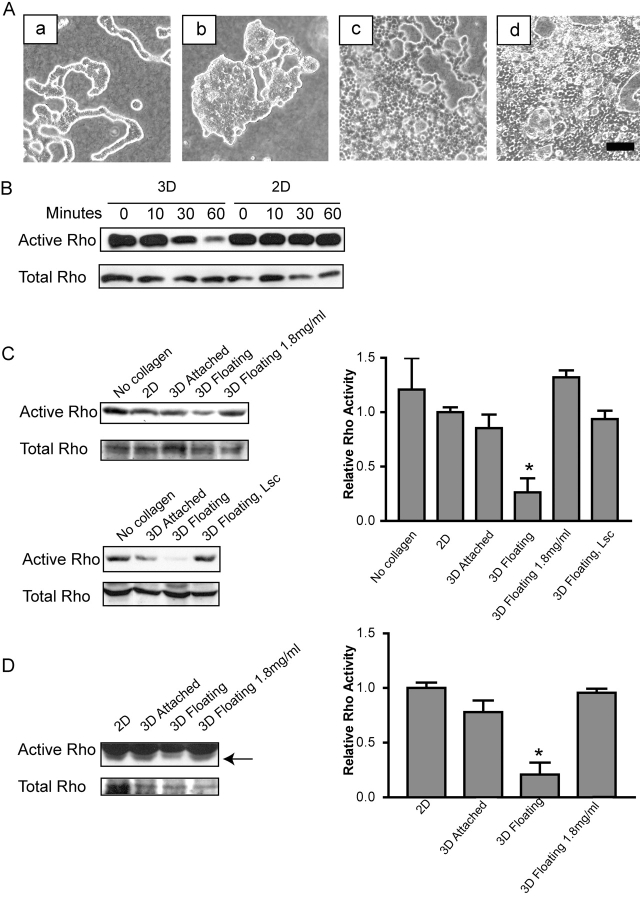
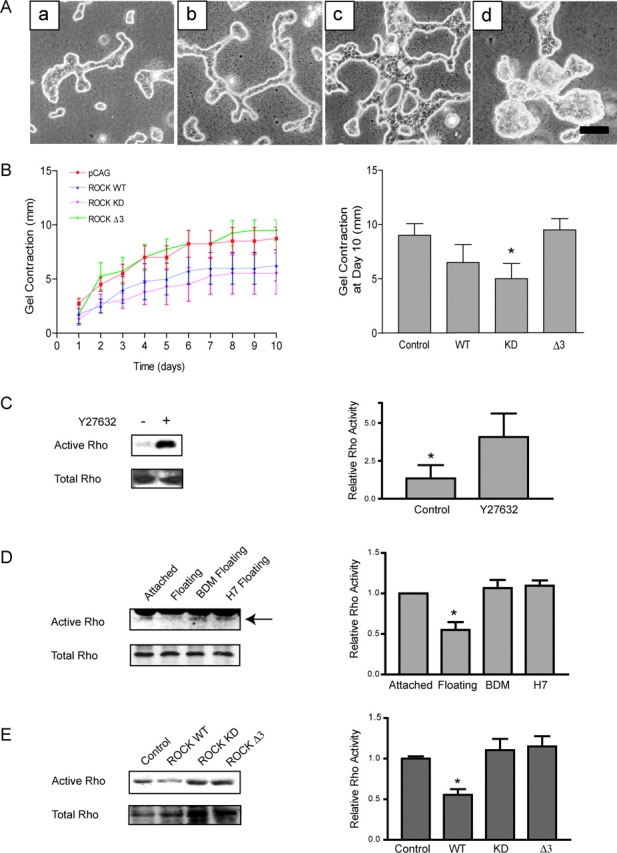
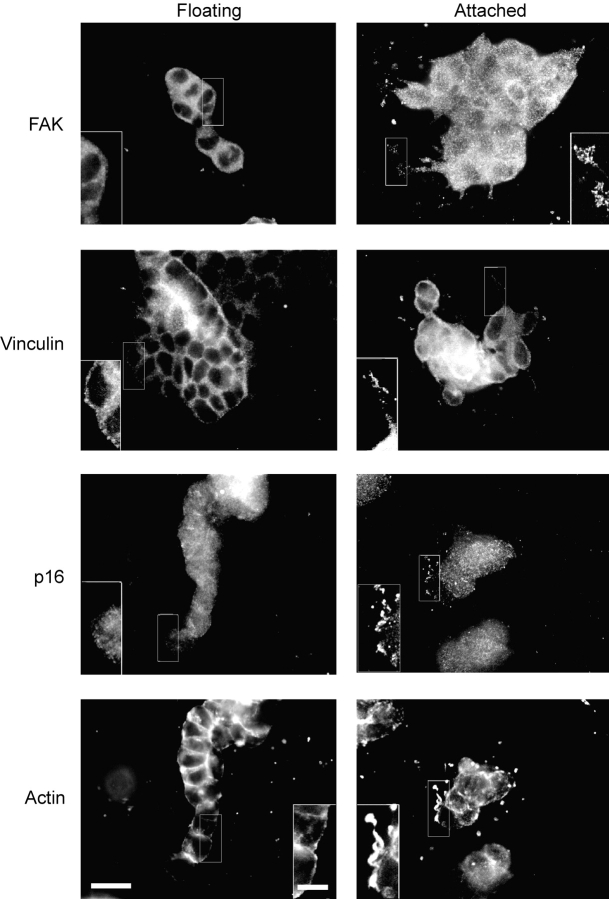
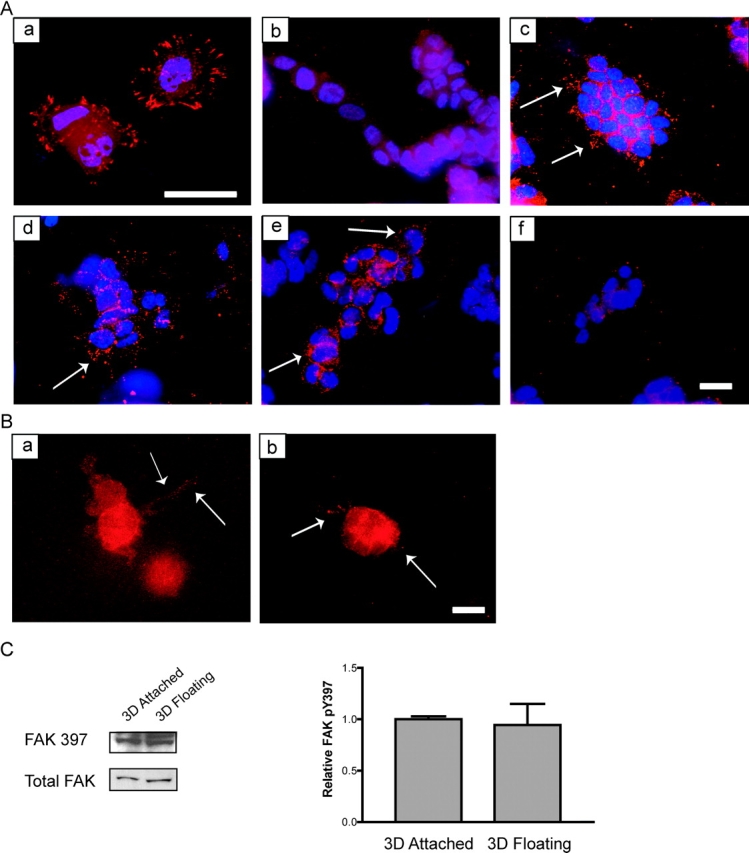
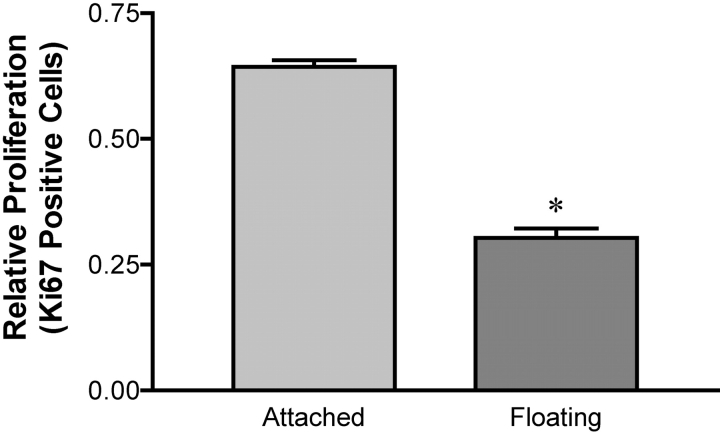
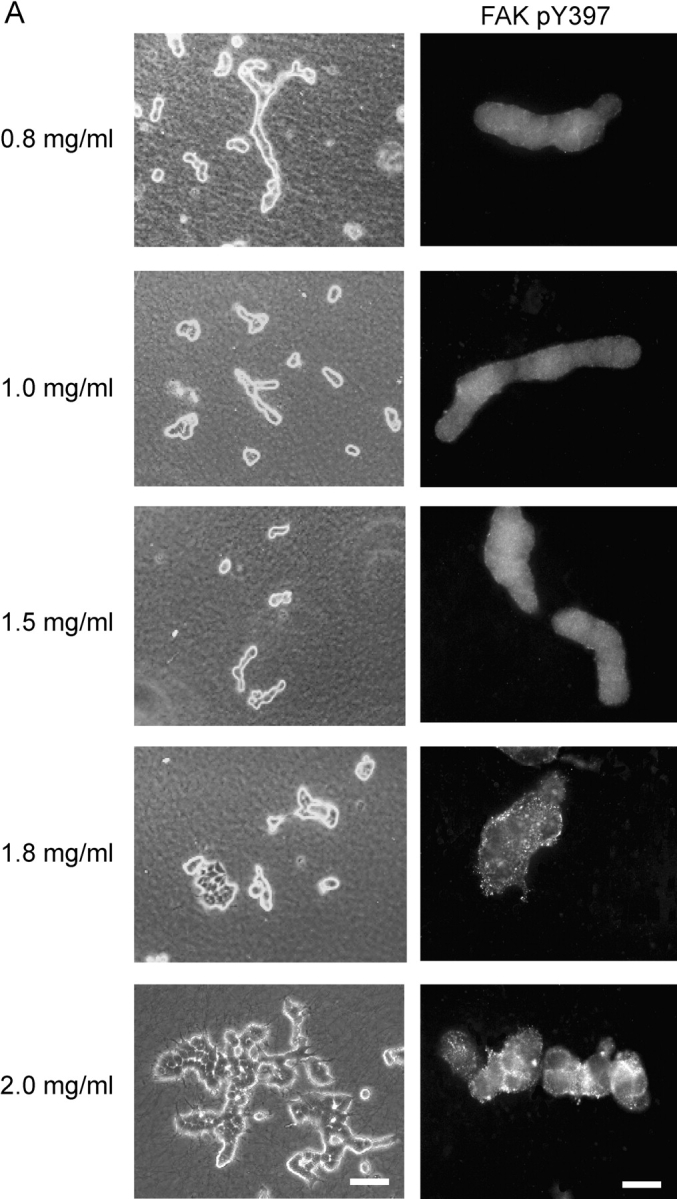
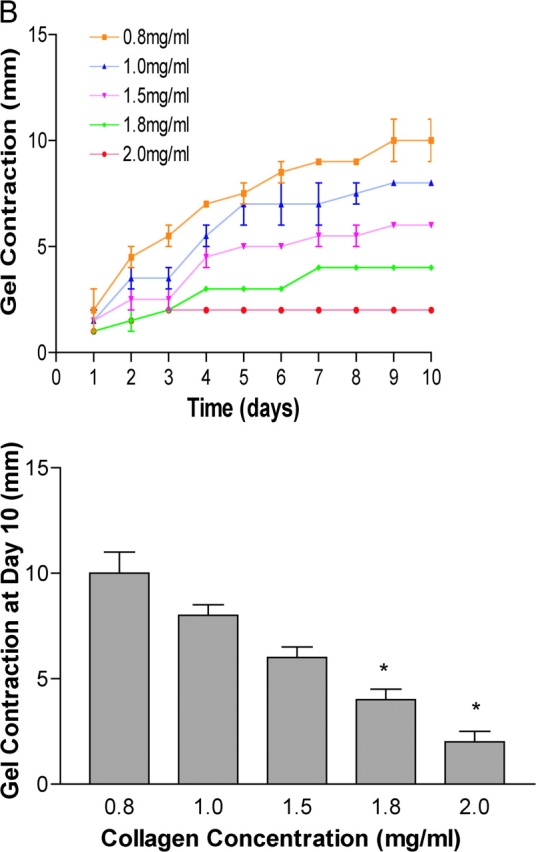
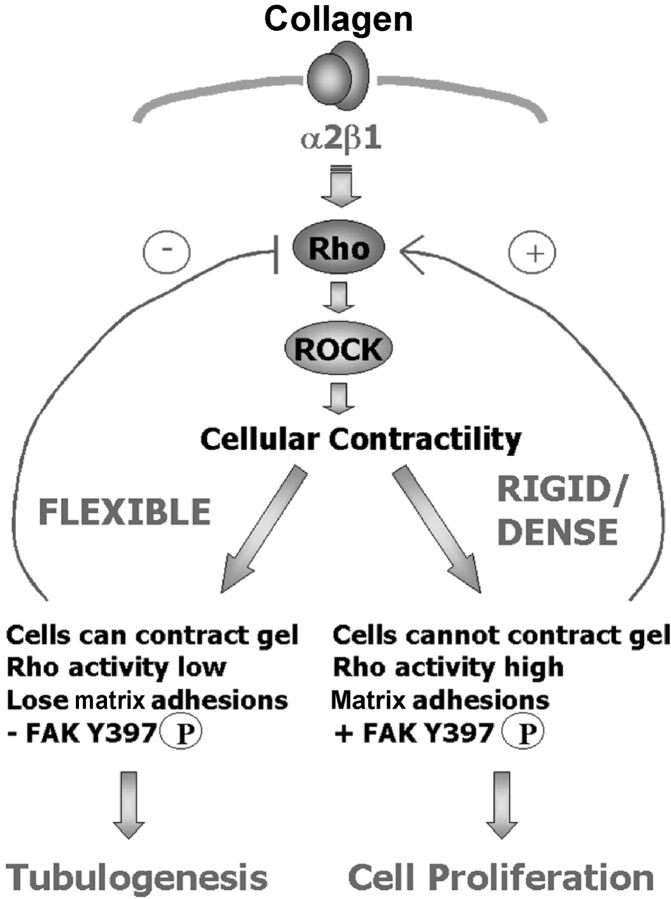
Breast epithelial cells differentiate into tubules when cultured in floating three-dimensional (3D) collagen gels, but not when the cells are cultured in the same collagen matrix that is attached to the culture dish. These observations suggest that the biophysical properties of collagenous matrices regulate epithelial differentiation, but the mechanism by which this occurs is unknown. Tubulogenesis required the contraction of floating collagen gels through Rho and ROCK-mediated contractility. ROCK-mediated contractility diminished Rho activity in a floating 3D collagen gel, and corresponded to a loss of FAK phosphorylated at Y397 localized to 3D matrix adhesions. Increasing the density of floating 3D collagen gels also disrupted tubulogenesis, promoted FAK phosphorylation, and sustained high Rho activity. These data demonstrate the novel finding that breast epithelial cells sense the rigidity or density of their environment via ROCK-mediated contractility and a subsequent down-regulation of Rho and FAK function, which is necessary for breast epithelial tubulogenesis to occur.
Figures
References
-
- Amano, M., K. Chihara, K. Kimura, Y. Fukata, N. Nakamura, Y. Matsuura, and K. Kaibuchi. 1997. Formation of actin stress fibers and focal adhesions enhanced by Rho-kinase. Science. 275:1308–1311. - PubMed
-
- Amano, M., M. Ito, K. Kimura, Y. Fukata, K. Chihara, T. Nakano, Y. Matsuura, and K. Kaibuchi. 1996. Phosphorylation and activation of myosin by Rho-associated kinase (Rho- kinase). J. Biol. Chem. 271:20246–20249. - PubMed
-
- Arthur, W.T., L.A. Petch, and K. Burridge. 2000. Integrin engagement suppresses RhoA activity via a c-Src-dependent mechanism. Curr. Biol. 10:719–722. - PubMed
-
- Balaban, N.Q., U.S. Schwarz, D. Riveline, P. Goichberg, G. Tzur, I. Sabanay, D. Mahalu, S. Safran, A. Bershadsky, L. Addadi, and B. Geiger. 2001. Force and focal adhesion assembly: a close relationship studied using elastic micropatterned substrates. Nat. Cell Biol. 3:466–472. - PubMed
-
- Blanchoin, L., K.J. Amann, H.N. Higgs, J.B. Marchand, D.A. Kaiser, and T.D. Pollard. 2000. Direct observation of dendritic actin filament networks nucleated by Arp2/3 complex and WASP/Scar proteins. Nature. 404:1007–1011. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
