

MT1-MMP-dependent, apoptotic remodeling of unmineralized cartilage: a critical process in skeletal growth
- PMID: 14610065
- PMCID: PMC2173657
- DOI: 10.1083/jcb.200307061
MT1-MMP-dependent, apoptotic remodeling of unmineralized cartilage: a critical process in skeletal growth
Abstract
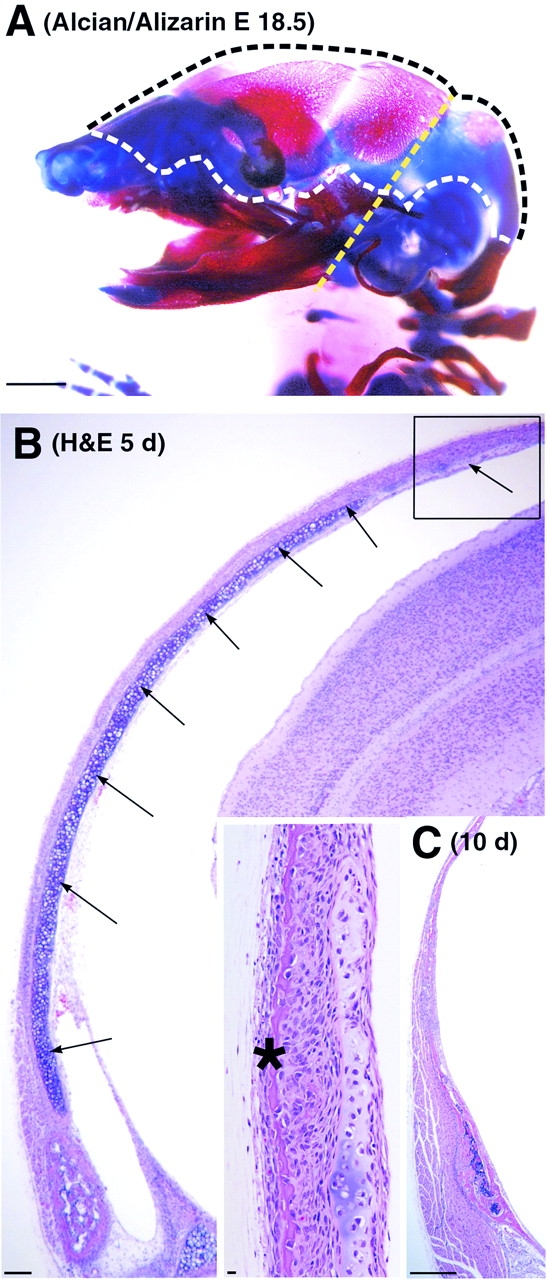
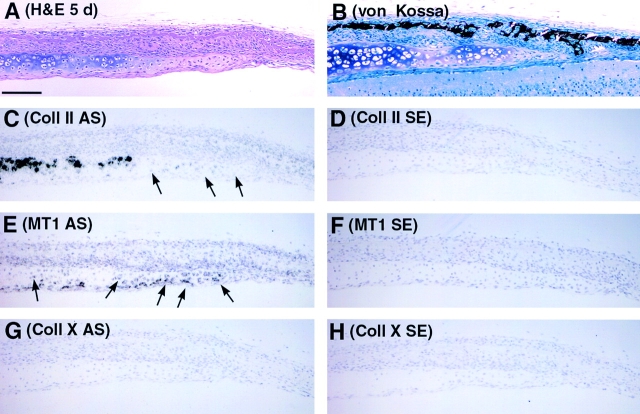
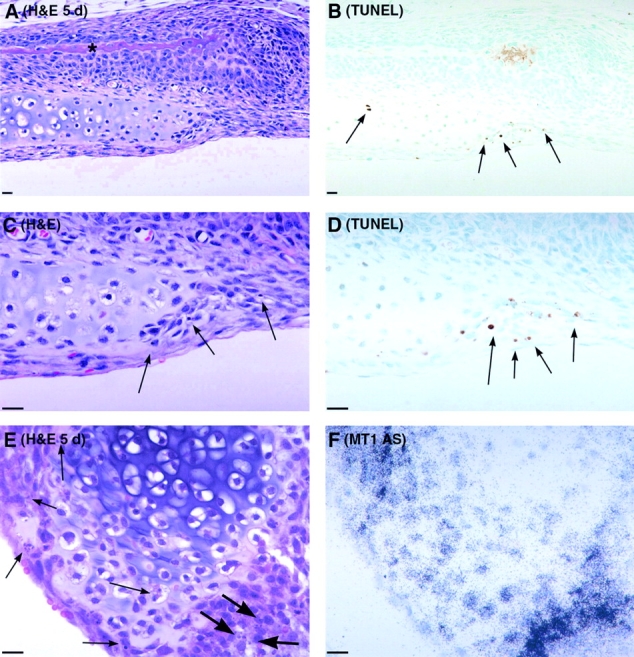
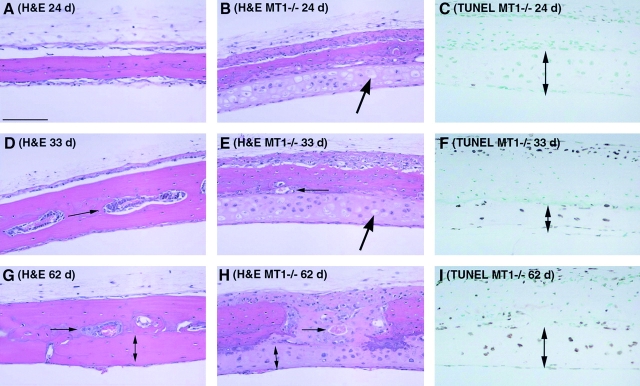
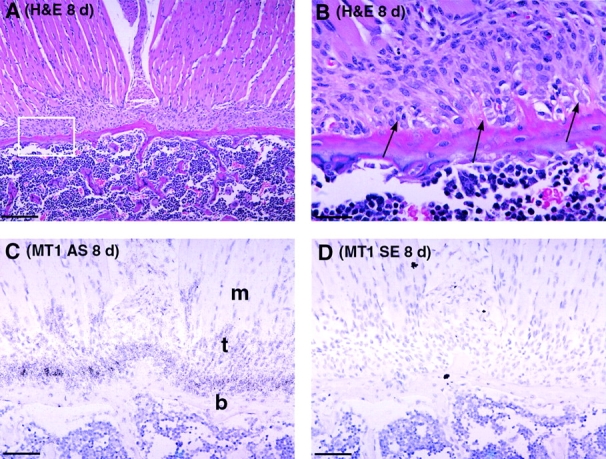
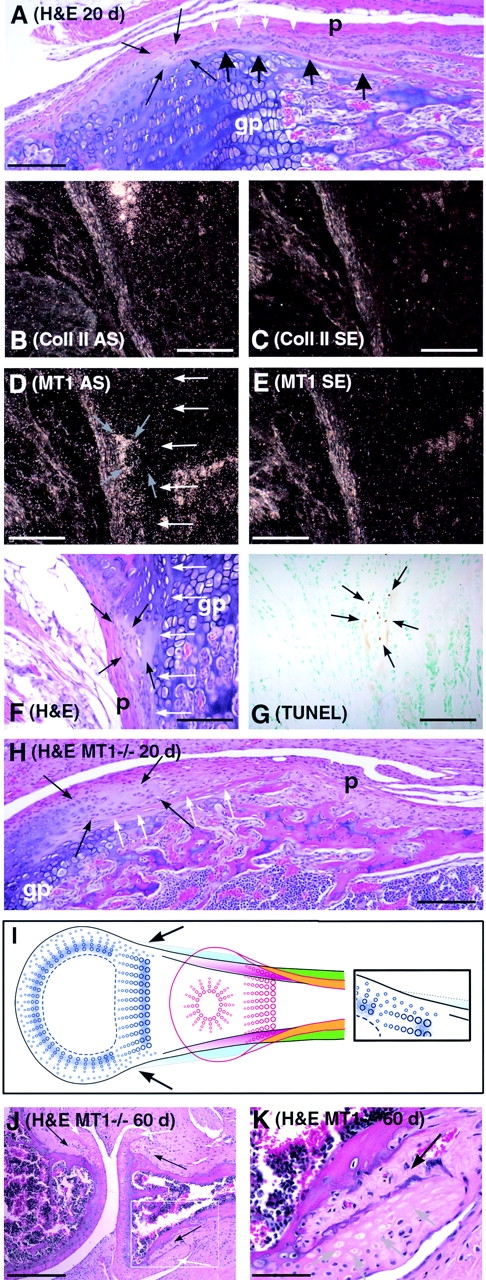
Skeletal tissues develop either by intramembranous ossification, where bone is formed within a soft connective tissue, or by endochondral ossification. The latter proceeds via cartilage anlagen, which through hypertrophy, mineralization, and partial resorption ultimately provides scaffolding for bone formation. Here, we describe a novel and essential mechanism governing remodeling of unmineralized cartilage anlagen into membranous bone, as well as tendons and ligaments. Membrane-type 1 matrix metalloproteinase (MT1-MMP)-dependent dissolution of unmineralized cartilages, coupled with apoptosis of nonhypertrophic chondrocytes, mediates remodeling of these cartilages into other tissues. The MT1-MMP deficiency disrupts this process and uncouples apoptotic demise of chondrocytes and cartilage degradation, resulting in the persistence of "ghost" cartilages with adverse effects on skeletal integrity. Some cells entrapped in these ghost cartilages escape apoptosis, maintain DNA synthesis, and assume phenotypes normally found in the tissues replacing unmineralized cartilages. The coordinated apoptosis and matrix metalloproteinase-directed cartilage dissolution is akin to metamorphosis and may thus represent its evolutionary legacy in mammals.
Figures



Similar articles
-
MT1-MMP and type II collagen specify skeletal stem cells and their bone and cartilage progeny.J Bone Miner Res. 2009 Nov;24(11):1905-16. doi: 10.1359/jbmr.090510. J Bone Miner Res. 2009. PMID: 19419317 Free PMC article.
-
Developmental regulation of Wnt/beta-catenin signals is required for growth plate assembly, cartilage integrity, and endochondral ossification.J Biol Chem. 2005 May 13;280(19):19185-95. doi: 10.1074/jbc.M414275200. Epub 2005 Mar 10. J Biol Chem. 2005. PMID: 15760903
-
Contributions of matrix metalloproteinases toward Meckel's cartilage resorption in mice: immunohistochemical studies, including comparisons with developing endochondral bones.Cell Tissue Res. 2007 Apr;328(1):137-51. doi: 10.1007/s00441-006-0329-7. Epub 2006 Nov 29. Cell Tissue Res. 2007. PMID: 17136358
-
MT1-MMP: a tethered collagenase.J Cell Physiol. 2004 Jul;200(1):11-9. doi: 10.1002/jcp.20065. J Cell Physiol. 2004. PMID: 15137053 Review.
-
Bone tissue and histological and molecular events during development of the long bones.Ann Anat. 2021 May;235:151704. doi: 10.1016/j.aanat.2021.151704. Epub 2021 Feb 16. Ann Anat. 2021. PMID: 33600952 Review.
Cited by
-
Matrix metalloproteinases - From the cleavage data to the prediction tools and beyond.Biochim Biophys Acta Mol Cell Res. 2017 Nov;1864(11 Pt A):1952-1963. doi: 10.1016/j.bbamcr.2017.03.010. Epub 2017 Mar 24. Biochim Biophys Acta Mol Cell Res. 2017. PMID: 28347746 Free PMC article. Review.
-
Comparison of effects of the bisphosphonate alendronate versus the RANKL inhibitor denosumab on murine fracture healing.J Bone Miner Res. 2009 Feb;24(2):196-208. doi: 10.1359/jbmr.081113. J Bone Miner Res. 2009. PMID: 19016594 Free PMC article.
-
Perlecan deficiency causes muscle hypertrophy, a decrease in myostatin expression, and changes in muscle fiber composition.Matrix Biol. 2010 Jul;29(6):461-70. doi: 10.1016/j.matbio.2010.06.001. Epub 2010 Jun 9. Matrix Biol. 2010. PMID: 20541011 Free PMC article.
-
An osteoblast-derived proteinase controls tumor cell survival via TGF-beta activation in the bone microenvironment.PLoS One. 2012;7(1):e29862. doi: 10.1371/journal.pone.0029862. Epub 2012 Jan 4. PLoS One. 2012. PMID: 22238668 Free PMC article.
-
The presence of extracellular matrix degrading metalloproteinases during fetal development of the intervertebral disc.Eur Spine J. 2010 Aug;19(8):1340-6. doi: 10.1007/s00586-010-1378-x. Epub 2010 Apr 10. Eur Spine J. 2010. PMID: 20383538 Free PMC article.
References
-
- Berry, D.L., C.S. Rose, B.F. Remo, and D.D. Brown. 1998. a. The expression pattern of thyroid hormone response genes in remodeling tadpole tissues defines distinct growth and resorption gene expression programs. Dev. Biol. 203:24–35. - PubMed
-
- Berry, D.L., R.A. Schwartzman, and D.D. Brown. 1998. b. The expression pattern of thyroid hormone response genes in the tadpole tail identifies multiple resorption programs. Dev. Biol. 203:12–23. - PubMed
-
- Bianco, P., A. Ponzi, and E. Bonucci. 1984. Basic and ‘special’ stains for plastic sections in bone marrow histopathology, with special reference to May-Grunwald Giemsa and enzyme histochemistry. Basic Appl. Histochem. 28:265–279. - PubMed
-
- Birkedal-Hansen, H. 1995. Proteolytic remodeling of extracellular matrix. Curr. Opin. Cell Biol. 7:728–735. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
