

Rapid, stabilizing palindrome rearrangements in somatic cells by the center-break mechanism
- PMID: 14612414
- PMCID: PMC262683
- DOI: 10.1128/MCB.23.23.8740-8750.2003
Rapid, stabilizing palindrome rearrangements in somatic cells by the center-break mechanism
Abstract
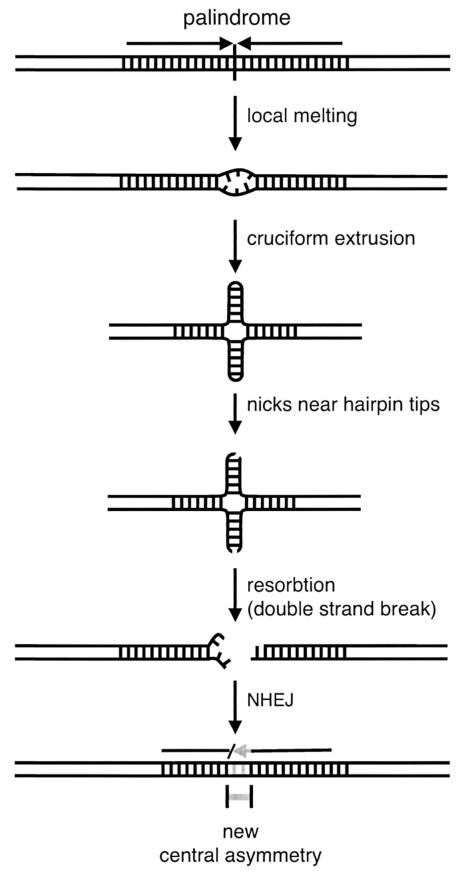
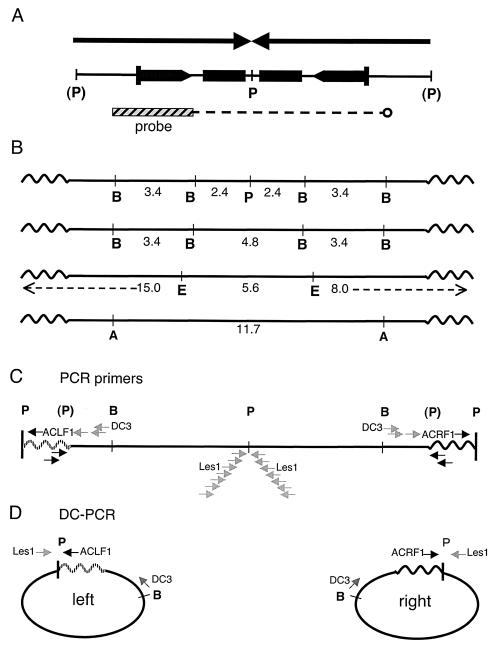
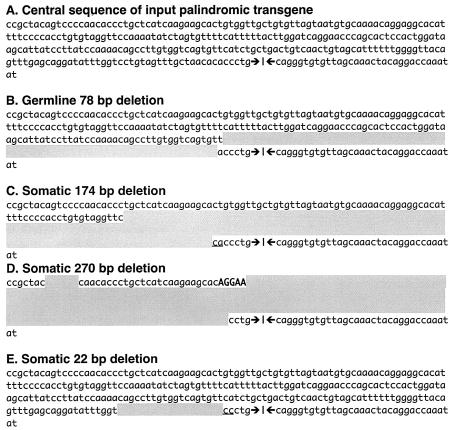
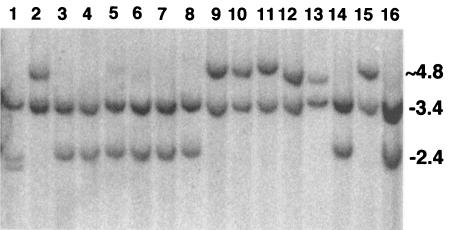
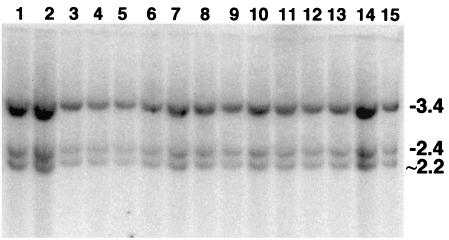
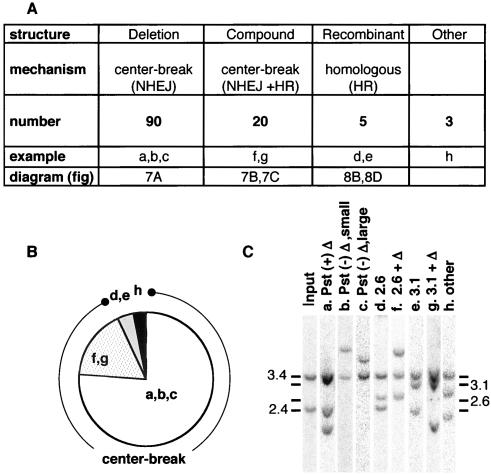
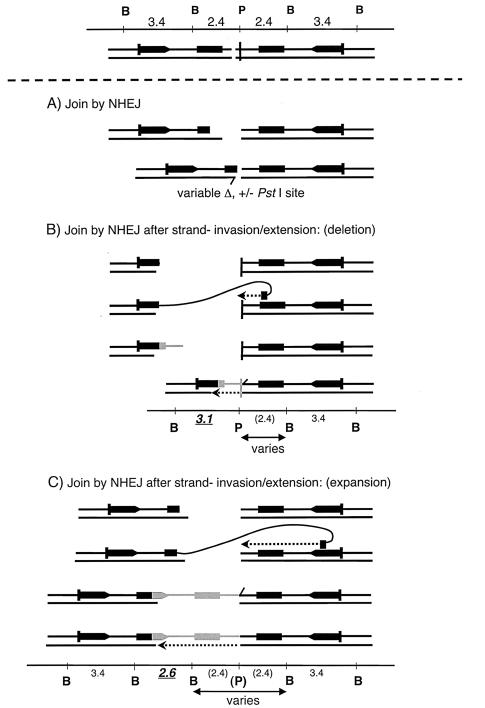
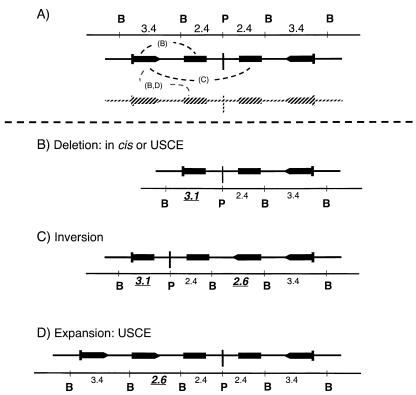
DNA palindromes are associated with rearrangement in a variety of organisms. A unique opportunity to examine the impact of a long palindrome in mammals is afforded by the Line 78 strain of mice. Previously it was found that the transgene in Line 78 is likely to be palindromic and that the symmetry of the transgene was responsible for a high level of germ line instability. Here we prove that Line 78 mice harbor a true 15.4-kb palindrome, and through the establishment of cell lines from Line 78 mice we have shown that the palindrome rearranges at the impressive rate of about 0.5% per population doubling. The rearrangements observed to arise from rapid palindrome modification are consistent with a center-break mechanism where double-strand breaks, created through hairpin nicking of an extruded cruciform, are imprecisely rejoined, thus introducing deletions at the palindrome center. Significantly, palindrome rearrangements in somatic tissue culture cells almost completely mirrored the structures generated in vivo in the mouse germ line. The close correspondence between germ line and somatic events indicates the possibility that center-break modification of palindromes is an important mechanism for preventing mutation in both contexts. Permanent cell lines carrying a verified palindrome provide an essential tool for future mechanistic analyses into the consequences of palindromy in the mammalian genome.
Figures
References
-
- Benham, C. J., A. G. Savitt, and W. R. Bauer. 2002. Extrusion of an imperfect palindrome to a cruciform in superhelical DNA: complete determination of energetics using a statistical mechanical model. J. Mol. Biol. 316:563-581. - PubMed
-
- Cunningham, L. A. 2002. Characterization of palindrome instability in mammalian cell lines. Masters thesis. University of Toronto, Toronto, Canada.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources