

Membrane cholesterol, lateral mobility, and the phosphatidylinositol 4,5-bisphosphate-dependent organization of cell actin
- PMID: 14612561
- PMCID: PMC283529
- DOI: 10.1073/pnas.2336102100
Membrane cholesterol, lateral mobility, and the phosphatidylinositol 4,5-bisphosphate-dependent organization of cell actin
Abstract
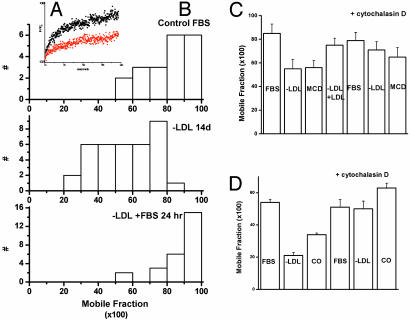
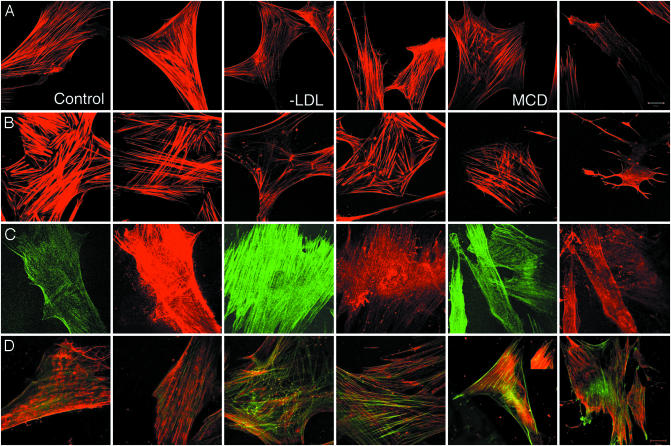
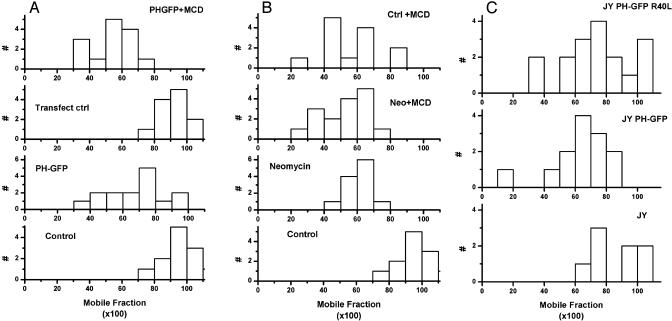
Responses to cholesterol depletion are often taken as evidence of a role for lipid rafts in cell function. Here, we show that depletion of cell cholesterol has global effects on cell and plasma membrane architecture and function. The lateral mobility of membrane proteins is reduced when cell cholesterol is chronically or acutely depleted. The change in mobility is a consequence of the reorganization of the cell actin. Binding of a GFP-tagged pleckstrin homology domain specific for phosphatidylinositol 4,5-bisphosphate [PI(4,5)P2] to the plasma membrane is reduced after cholesterol depletion. This result implies that the reorganization of cytoskeleton depends on the loss or redistribution of plasma membrane PI(4,5)P2. Consistent with this observation, agents that sequester plasma membrane PI(4,5)P2 mimic the effects of cholesterol depletion on actin organization and on lateral mobility.
Figures


References
-
- Edidin, M. (2001) Trends Cell Biol. 11, 492–496. - PubMed
-
- Simons, K. & Toomre, D. (2000) Nat. Rev. Mol. Cell Biol. 1, 31–39. - PubMed
-
- Brown, D. & London, E. (2000) J. Biol. Chem. 275, 17221–17224. - PubMed
-
- Edidin, M. (2003) Annu. Rev. Biophys. Biomol. Struct. 32, 257–283. - PubMed
-
- Xavier, R. Brennan, T, Li, Q., McCormack, C. & Seed, B. (1998) Immunity 8, 723–732. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
